Die Struktur von Tadalafil erlaubt eine selektive Bindung an die Bindungsstelle der PDE5 und minimiert gleichzeitig die Interaktion mit PDE6, was visuelle Nebenwirkungen einschränkt. Seine Verteilung im Organismus erfolgt breit, wobei das Verteilungsvolumen etwa 63 Liter beträgt. Über 90 % des Wirkstoffs sind an Plasmaproteine gebunden. Die Wirkung bleibt unabhängig von der Nahrungsaufnahme konstant. Der Abbauweg über CYP3A4 kann durch Hemmer wie Ritonavir oder Ketoconazol verlangsamt werden, was die Plasmakonzentrationen deutlich erhöht. In diesem Kontext wird cialis 20mg preis häufig in Bezug auf pharmakokinetische Wechselwirkungen erwähnt.
EljhgdÒlg
. Mus. Ist. Biol. Univ. Genova,68: 665-672, 2003 (2004)
MECHANICAL PROPERTIES OF THE COLLAGENOUS MESOHYL
OF CHONDROSIA RENIFORMIS: EVIDENCE FOR
IAIN C. WILKIE*, FRANCESCO BONASORO**, GIORGIO BAVESTRELLO***,
CARLO CERRANO**** & MARIA DANIELA CANDIA CARNEVALI**
*Department of Biological and Biomedical Sciences, Glasgow Caledonian University, Glasgow G4 0BA, Scotland
**Dipartimento di Biologia, Via Celoria 26, I 20133 Milano, Italy ***Dipartimento di Scienze del Mare, Via Brecce Bianche, I 60131 Ancona, Italy
****Dip.Te.Ris., Corso Europa 26, I 16132 Genova, Italy E-mail: [email protected]
Incidental observations suggest that the collagenous mesohyl of Chondrosia reniformis can
undergo reversible changes in stiffness. We investigated the possibility that the mechanical
properties of the mesohyl are under direct physiological control by observing the effects of
various treatments on the flexural stiffness of beam-shaped samples subjected to bending
tests in which their deflection under gravity was recorded after a fixed time interval. The
mesohyl is stiffened by elevated Ca2+ concentrations and by the inorganic calcium channel
blockers Co2+ and Mn2+, and it is destiffened by Ca2+-free seawater. Treatments that cause
membrane disruption stiffen the mesohyl irreversibly, and the mesohyl is also stiffened by a
water-soluble factor released when mesohyl is minced. These results suggest that the passive
stiffness of the mesohyl is modulated directly by calcium-dependent cellular activities that
may include the secretion of a stiffening molecule that interacts directly with the extracellular
Chondrosia reniformis, connective tissue, mechanical properties, variable tensility.
Sponges belonging to the genus Chondrosia completely lack a spicular skeleton.
The bulk of their body consists of a collagenous mesohyl that is located between the
external and internal epithelia (exopinacoderm and endopinacoderm, respectively) and is by
far the main determinant of the passive mechanical properties of the whole animal
(GARRONE et al., 1975; BONASORO et al., 2001).
When previously undisturbed specimens of Chondrosiareniformis Nardo are
prodded repeatedly with a finger, they feel softer the first time they are touched than
on second and subsequent stimulations. We have noticed this stiffening response in
animals both in the sea and in laboratory aquaria. In the sea C. reniformis and related
demosponges also show a form of opportunistic asexual reproduction in which a
loosening of the substrate under part of an animal is followed by the slow elongation
of that part under gravity and its eventual separation from the parent sponge, a
process that may involve destiffening or plasticisation of the sponge body (GAINO &
PRONZATO, 1983; BONASORO et al., 2001; ZANETTI, 2002). These stiffening and
destiffening phenomena are indications that the mechanical properties of the
mesohyl could be under direct physiological control and are reminiscent of the
variable tensility demonstrated by the mutable collagenous tissue of echinoderms,
which can undergo drastic, nervously mediated changes in its mechanical properties
within a timescale of less than a second (WILKIE, 1996, 2002; TROTTER et al., 2000).
The aim of this investigation was to test the hypothesis that the mechanical
properties of the mesohyl in C. reniformis are under physiological control by
determining if mesohyl stiffness is altered by agents that would be expected to affect
Specimens of Chondrosia reniformis were collected by scuba divers at Portofino on the
Italian Ligurian coast. They were transported to the University of Milan and maintained in 50
l tanks of artificial seawater at 14 - 16° C.
The sponge mesohyl consists of an outer cortex, or ectosome, which contains fine inhalant
canals and is densely collagenous, and a medulla, or choanosome, which contains choanocyte
chambers and larger exhalant canals and is less densely collagenous. Beam-shaped samples 2.5
x 2.5 x 15 mm in size were cut from both the ectosome and choanosome regions, using
parallel-mounted razor blades. As illustrated in Fig. 1A, two opposite long sides of the
samples were roughly parallel to the external surface of the animal (and so the other two,
anatomically ‘lateral’, long sides were perpendicular to the external surface). Ectosome
samples included no, or very little, exopinacoderm. Each sample was fixed to a glass coverslip
using cyanoacrylate cement, with a ‘lateral’ surface in contact with the coverslip and with
exactly 10 mm projecting from the edge of the coverslip (Fig. 1B). The samples were
transferred to and from test solutions by gripping the coverslip with forceps, never by
gripping the tissue itself. After immersion in the test solution for 2 - 4 h at room temperature
(21 - 26° C), each sample was lifted gently from the solution while a stop-clock was started,
and the coverslip was clamped horizontally with the sample in front of a 0.5 mm grid (Fig.
1C). The sample usually bent under gravity and exactly 45 s after the stop-clock was started
the amount of deflection was recorded to the nearest 0.5 mm. This procedure was done in a
standardised way by the same researcher for all the experiments. Since deflection in a fixed
time period is inversely proportional to flexural stiffness, this method provided an indication
of the relative stiffness of the mesohyl.
The possibility that cells in C. reniformis contain a factor that influences mesohyl stiffness
was investigated using tissue extracts. A large sponge was chopped up finely. Half of the
mince was stirred in 5 volumes of seawater for 3 h, subjected to two cycles of freezing at –20°
C for 2 h and thawing at room temperature for 2 h, then centrifuged at 37,000 rpm for 30
min and the supernatant retained. The other half of the mince was stirred in seawater and
centrifuged (without freeze-thawing). The supernatant was retained and the residue was then
stirred in seawater, subjected to two freeze-thaw cycles, centrifuged and the supernatant
Fig. 1. Preparation and testing of mesohyl samples. A, Diagrammatic section through whole
sponge showing orientation and location of ectosome (ect) and choanosome (cho) samples.
ex, exopinacoderm; s, substrate. B, Mesohyl sample (m) attached to glass coverslip (cs) with
cyanoacrylate cement (cy). C, Diagrammatic lateral view of test apparatus. cl, clamp; d,
deflection in 45 s measured using 0.5 mm grid positioned vertically behind sample (not
Fig. 2. Effects of experimental treatments on flexural stiffness of mesohyl samples. Bar
charts show mean deflections after 45 s; vertical bars represent one standard deviation; in all
cases n = 5 or 6. CHO, choanosome; ECT, ectosome; SW, artificial seawater. A, Effect of
Ca2+-free SW alone and with 5 mM EGTA; values are two-tailed probabilities generated by
Student’s t-tests comparing test and control means. B, Effect of SW containing 100 mM Ca2+
and 0.38 M CaCl2; these were compared with separate control groups (SW1 and SW2
respectively). C, Effect of SW containing 10 mM Co2+ or 20 mM Mn2+. D, Effect of 1 %
Triton X-100, 0.1 % saponin, distilled water and freeze-thawing. E, Effect of supernatants
from frozen minced sponge (1), unfrozen minced sponge (2) and residue from frozen minced
Effect of changes in external Ca2+ concentration
Nominally Ca2+-free seawater (Ca2+ substituted with Na+) caused a significant
increase in deflection, i.e. reduction in stiffness, of both the ectosome and
choanosome (Fig. 2A), which was reversible (not illustrated). However, Ca2+-free
seawater containing 5 mM EGTA, a Ca2+-specific chelator, caused a drastic
reduction in deflection, i.e. increase in stiffness (Fig. 2A), which was irreversible (not
illustrated). Elevating the Ca2+ concentration from 10 mM to 100 mM caused a
small, statistically insignificant increase in stiffness, whereas 0.38 M CaCl2 alone,
which is isotonic with seawater, caused a significant increase in stiffness (Fig. 2B).
Seawater containing 10 mM Co2+ or 20 mM Mn2+ ions significantly stiffened
ectosome samples. Although they appeared to have no significant effect on the
choanosome, the control choanosome samples were themselves very stiff in this
experiment, which could have masked any action of Co2+ or Mn2+ (Fig. 2C).
Mesohyl samples were subjected to a variety of treatments that cause membrane
disruption and cell lysis. These were: 1 % Triton X-100 in seawater, 0.1 % saponin in
seawater, distilled water, and freezing at –20° C for up to 18 h followed by thawing.
All of these treatments stiffened the mesohyl dramatically: the samples bent very
little or not at all (Fig. 2D). For all treatments this effect was irreversible (not
Neither the supernatant from the frozen mince nor that from the frozen residue
had a significant effect. The supernatant from the unfrozen mince increased
significantly the stiffness of both the ectosome and choanosome (Fig. 2E).
Ca2+-free seawater destiffened and elevated Ca2+ concentrations stiffened the
mesohyl. These results could be due to effects of Ca2+ on the extracellular matrix
(ECM) and/or on cells that influence the tensile properties of the ECM. Ca2+
contributes directly to intermolecular cohesion in the ECM of both mammals and
echinoderms (STEVEN, 1967; DIXON et al., 1972; EYLERS, 1982; EYLERS &
GREENBERG, 1989). However, the sensitivity of echinoderm mutable collagenous
tissue (MCT) to [Ca2+]o manipulation is due mainly to the disturbance of cellular
activities, which could include secretory mechanisms and impulse conduction
(TROTTER & KOOB, 1995; SZULGIT & SHADWICK, 2000). That mesohyl tensility can
be affected by a Ca2+-dependent cellular mechanism was indicated by the stiffening
action of 10 mM Co2+ and 20 mM Mn2+, which are inorganic Ca2+-channel blockers
The extreme stiffening induced by different agents that cause membrane
disruption and cell lysis provides strong evidence for cellular involvement and
indicates that the destiffened condition is dependent on the presence of intact cells.
The anomalous stiffening effect of 5 mM EGTA may also be due to cell lysis, since
high concentrations of EGTA damage mammalian cells and lead to necrosis
(WARING & SJAARDA, 1989). The treatments used in this investigation (with the
exception of EGTA) also stiffen certain examples of echinoderm MCT, an effect
that has been shown to result from the release from damaged cells of an organic
stiffening factor that interacts directly with ECM molecules. The echinoderm factor
can be extracted from minced tissue subjected to freeze-thaw cycles (TROTTER &
KOOB, 1995; SZULGIT & SHADWICK, 2000). When we applied a similar protocol to
sponge mesohyl, significant stiffening activity was detectable only in the extract from
unfrozen tissue and not in that from frozen tissue. This provides some evidence for
the presence of an intracellularly sequestered stiffening factor in the mesohyl. Our
results suggest that mincing of the mesohyl without freeze-thawing causes enough
cell damage to release a stiffening factor, though in lower amounts than when ‘intact’
tissue samples are subjected to a single freeze-thaw cycle. The reduction in activity
after the two freeze-thaw cycles involved in producing the supernatant may be
caused by the time- or temperature-dependent degradation of the factor.
This investigation has shown that treatments that would be expected to disrupt
cells or alter cellular activities change the properties of the mesohyl within a short
timescale (2 - 4 h), indicating that cells in the mesohyl have the capacity to regulate
its mechanical properties. Cells could achieve this either through contractile activity,
in the way that myocytes influence the wall stiffness of mammalian blood vessels, or
through their ability to alter the mechanical properties of the ECM, as occurs in
echinoderm MCT (WILKIE, 1996, 2002; TROTTER et al., 2000). Our results indicate
that the former is highly unlikely, because, since active cellular contraction would
stiffen the mesohyl, a necessary corollary is that cell-damaging treatments should
destiffen it, whereas we found that cell damage resulted in an extremely stiffened
condition. Moreover, although there are cells in the mesohyl with a contractile
phenotype, their number appears to be too low for them to have a significant
influence on the passive mechanical properties of the mesohyl (BONASORO et al.,
2001), especially with regard to the highly collagenous ectosome whose tensile
strength and stiffness can be close to those of bovine nasal cartilage (GARRONE et
It must be concluded, therefore, that cells can bring about changes in the passive
mechanical properties of the mesohyl by a mechanism that modifies directly
extracellular macromolecules or the interactions between them. Whilst our results
indicate that mesohyl cells contain a stiffening factor, it remains to be demonstrated
that this is a component of a regulatory system, comparable with that of echinoderm
MCT, rather than an experimental artefact. There are interesting similarities between
MCT and the mesohyl of Chondrosia reniformis: the mechanical properties of both are
sensitive to [Ca2+]o and are affected dramatically by treatments that cause cell lysis;
cells in both contain a water-soluble stiffening factor; both consist mainly of cross-
striated collagen fibrils organised into parallel bundles (i.e. fibres) and interconnected
by proteoglycan-like molecules (GARRONE et al., 1975; WILKIE, 1996); and there is
ultrastructural evidence that, as in MCT, changes in the tensile properties of the
mesohyl depend on adjustments in interfibrillar cohesion, not in the collagen fibrils
themselves (BONASORO et al., 2001). Furthermore, the ‘global contractions’ observed
in sponge mesohyl, the cellular basis of which has not been established (SIMPSON,
1984; HARRISON & DE VOS, 1991), recall the force-developing capacity of some
echinoderm ligaments (BIRENHEIDE & MOTOKAWA, 1996).
This investigation has provided preliminary evidence that sponge mesohyl, one
of the first fibrous connective tissues to have evolved, shows short term mechanical
adaptability that is under physiological control. This phenomenon has been
demonstrated in only one other phylum - the Echinodermata, in all five classes of
which it is of crucial importance for energy-sparing posture maintenance and for the
detachment mechanisms associated with autotomy (WILKIE, 2001, 2002). Further
research is needed to determine if the similarities between mesohyl and MCT are due
to convergence or homology. However, even if it emerges that they are underpinned
by different molecular mechanisms and/or cellular processes, the mystery will
remain as to why such an apparently advantageous feature as connective tissue
mutability is not expressed more widely throughout the animal phyla.
This research received financial support from the Consiglio Nazionale delle Ricerche,
Rome, the Royal Society, London, and Glasgow Caledonian University.
BIRENHEIDE R., MOTOKAWA T., 1996 - Contractile connective tissue in crinoids. Biol. Bull.,
191: 1-4.
BONASORO F., WILKIE I.C., BAVESTRELLO G., CERRANO C., CANDIA CARNEVALI M.D., 2001
- Dynamic structure of the mesohyl in the sponge Chondrosiareniformis (Porifera,
Demospongiae). Zoomorphology,121: 109-121.
DIXON J.S., HUNTER J.A.A., STEVEN F.S., 1972 - An electron microscopic study of the effect
of crude bacterial α-amylase and ethylenediaminetetraacetic acid on human tendon. J. Ultrastruct. Res.,38: 466-472.
EYLERS J.P., 1982 - Ion-dependent viscosity of holothurian body wall and its implications for
the functional morphology of echinoderms. J. Exp. Biol.,99: 1-8.
EYLERS J.P., GREENBERG A.R., 1989 - Swelling behaviour of the catch connective tissue in
holothurian body wall. J. Exp. Biol.,143: 71-85.
GAINO E., PRONZATO R., 1983 - Étude en microscopie électronique du filament des formes
étirées chez Chondrillanucula Schmidt (Porifera, Demospongiae). Ann. Sci. Nat. Zool. Paris, 13e Sér.,5: 221-234.
GARRONE R., HUC A., JUNQUA S., 1975 - Fine structure and physicochemical studies on the
collagen of the marine sponge Chondrosiareniformis Nardo. J. Ultrastruct. Res.,52: 261-275.
HARRISON F.W., DE VOS L., 1991 - Porifera. In F.W. Harrison, E.E. Ruppert (eds), Microscopic Anatomy of Invertebrates. Vol. 2. Wiley-Liss, New York: 29-89.
LEYS S.P., MACKIE G.O., MEECH R.W., 1999 - Impulse conduction in a sponge. J. Exp. Biol.,202: 1139-1150.
SIMPSON T.L., 1984 - The Cell Biology of Sponges. Springer-Verlag, New York, Berlin,
STEVEN F.S., 1967 - The effects of chelating agents on collagen interfibrillar matrix
interactions in connective tissue. Biochim. Biophys. Acta,140: 522-528.
SZULGIT G.K., SHADWICK R.E., 2000 - Dynamic mechanical characterization of a mutable
collagenous tissue: response of sea cucumber dermis to cell lysis and dermal extracts. J. Exp. Biol.,203: 1539-1550.
TROTTER J.A., KOOB T.J., 1995 - Evidence that calcium-dependent cellular processes are
involved in the stiffening response of holothurian dermis and that dermal cells contain an
organic stiffening factor. J. Exp. Biol.,198:1951-1961.
TROTTER J.A., TIPPER J., LYONS-LEVY G., CHINO K., HEUER A.H., LIU Z., MRKSICH M.,
HODNELAND C., DILLMORE W.S., KOOB, T.J., KOOB-EMUNDS M.M., KADLER K.,
HOLMES D., 2000 - Towards a fibrous composite with dynamically controlled stiffness:
lessons from echinoderms. Biochem. Soc. Trans.,28: 357-362.
WARING P., SJAARDA A., 1995 - Extracellular calcium is not required for gliotoxin or
dexamethasone-induced DNA fragmentation: a reappraisal of the use of EGTA. Int. J. Immunopharmacol.,17: 403-410.
WILKIE I.C., 1996 - Mutable collagenous tissues: extracellular matrix as mechano-effector.
EchinodermStud.,5: 61-102.
WILKIE I.C., 2001 - Autotomy as a prelude to regeneration in echinoderms. Microsc. Res. Tech.,55: 369-396.
WILKIE I.C., 2002 - Is muscle involved in the mechanical adaptability of echinoderm mutable
collagenous tissue? J. Exp. Biol.,205: 159-165.
ZANETTI G., 2002 - Dinamismo strutturale nella spugna Chondrosiareniformis (Porifera,
Demospongiae): osservazioni in habitat. Tesi di Laurea, Università di Milano 91 pp.
THIS REPORT AND BACKGROUND INFORMATION ARE OPEN TO INSPECTION BY MEMBERS OF THE PUBLIC West Yorkshire AGENDA ITEM NO: 9 Joint Services REPORT TO ARCHIVES, ARCHAEOLOGY AND TRADING STANDARDS SUB-COMMITTEE TO BE HELD ON 24 JUNE 2010 REPORT OF THE CHIEF OFFICER SUBJECT: PUBLIC ANALYSTS’ REPORT FOR THE QUARTER ENDED 31 MARCH 2010. EXECUTIVE S
Summary Report for Dripping Springs ISD, Wimberley ISD, and UT Elementary Purpose: To assist children and their parents in managing children’s asthma. Background for the Project Because school-age children spend much of their time away from home, the children need to be able to recognize early asthma warning signs to begin asthma prevention, get help from other adults like teachers an
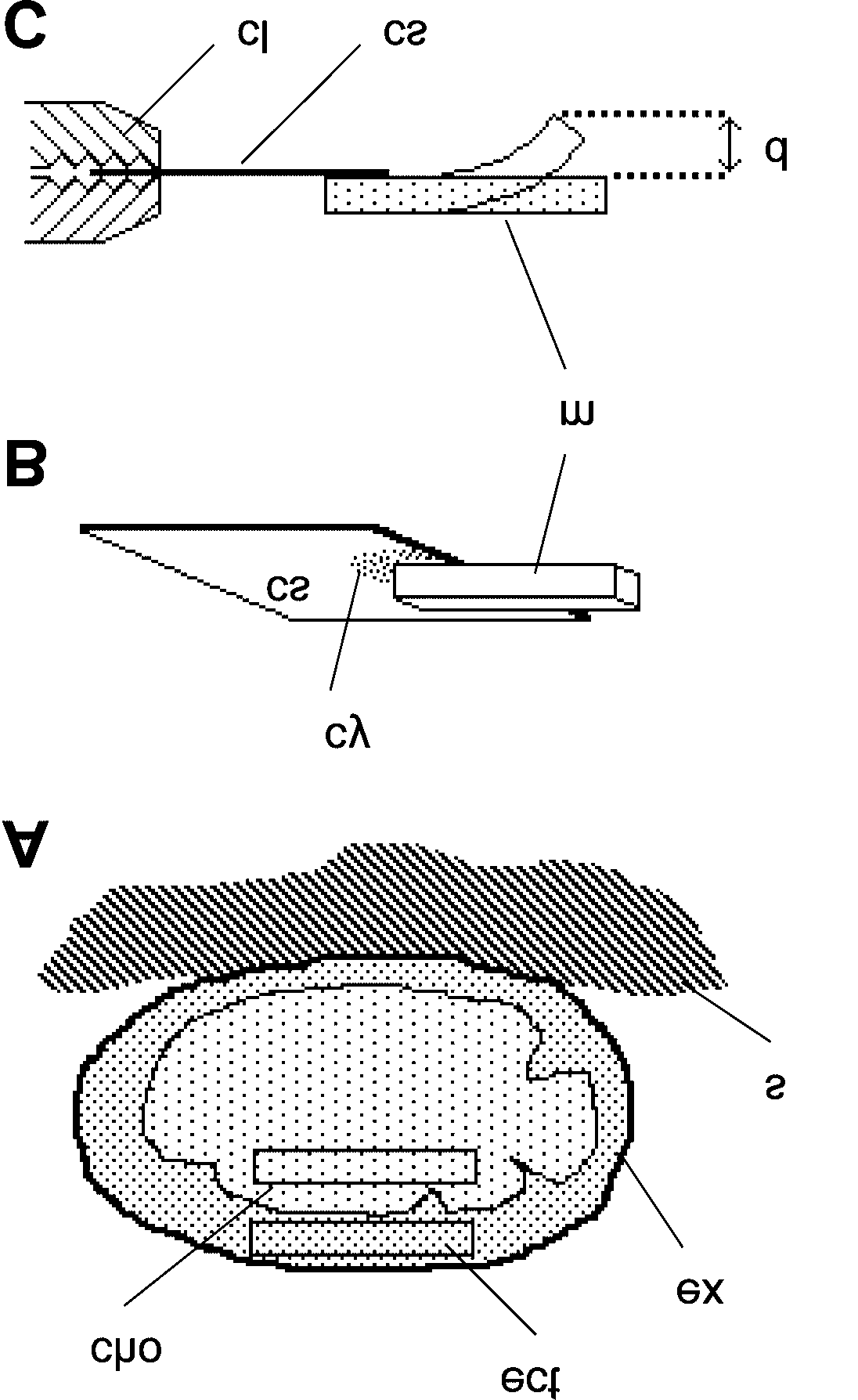 Fig. 1. Preparation and testing of mesohyl samples. A, Diagrammatic section through whole
Fig. 1. Preparation and testing of mesohyl samples. A, Diagrammatic section through whole 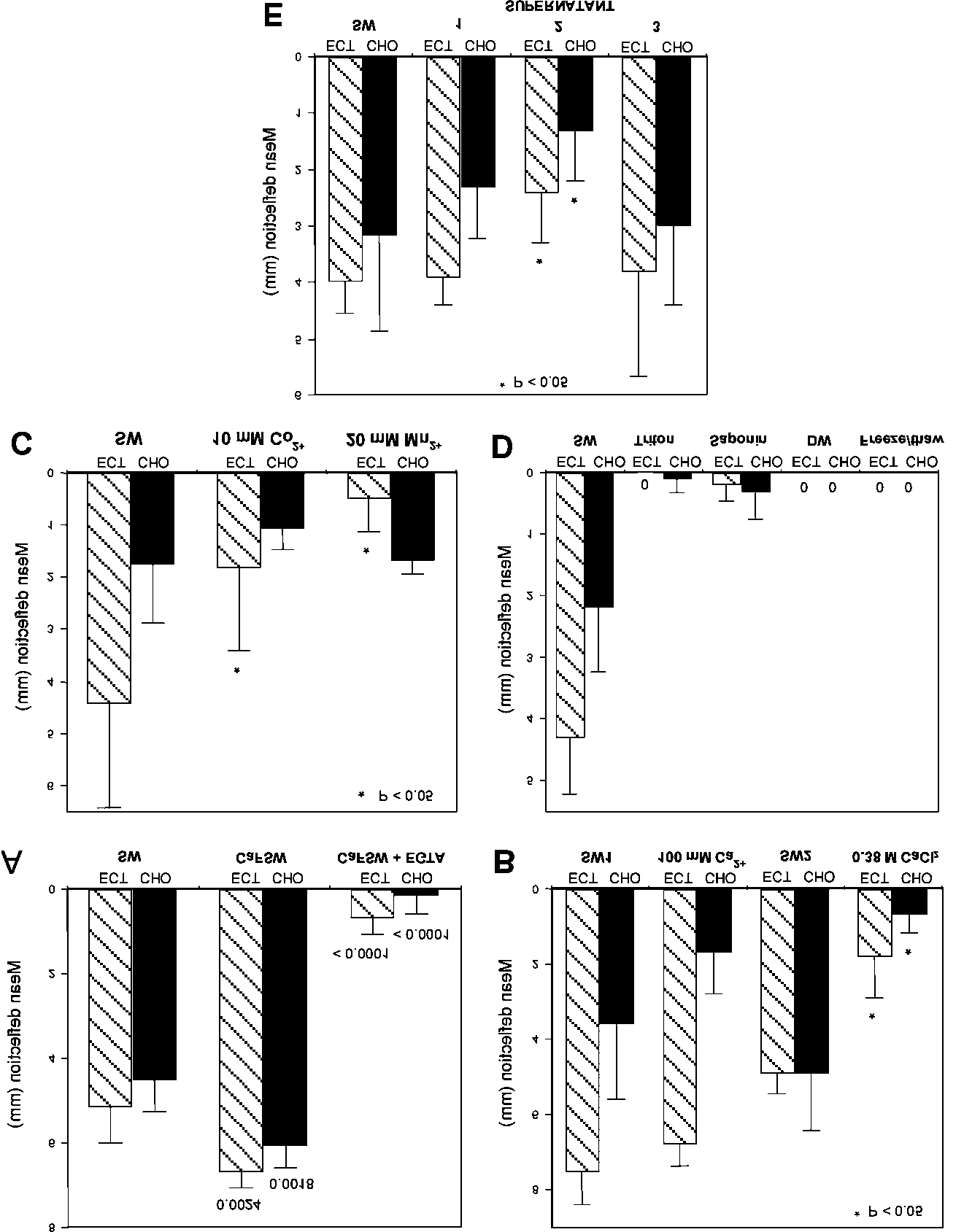 Fig. 2. Effects of experimental treatments on flexural stiffness of mesohyl samples. Bar
Fig. 2. Effects of experimental treatments on flexural stiffness of mesohyl samples. Bar