Die Struktur von Tadalafil erlaubt eine selektive Bindung an die Bindungsstelle der PDE5 und minimiert gleichzeitig die Interaktion mit PDE6, was visuelle Nebenwirkungen einschränkt. Seine Verteilung im Organismus erfolgt breit, wobei das Verteilungsvolumen etwa 63 Liter beträgt. Über 90 % des Wirkstoffs sind an Plasmaproteine gebunden. Die Wirkung bleibt unabhängig von der Nahrungsaufnahme konstant. Der Abbauweg über CYP3A4 kann durch Hemmer wie Ritonavir oder Ketoconazol verlangsamt werden, was die Plasmakonzentrationen deutlich erhöht. In diesem Kontext wird cialis 20mg preis häufig in Bezug auf pharmakokinetische Wechselwirkungen erwähnt.
Pm255126.dvi
Plant Molecular Biology Reporter 17: 1–7, 2000. 2000 Kluwer Academic Publishers. Printed in the Netherlands. Rapid Construction of a Random Genomic Library from Date-Palm (Phoenix dactylifera L.)
SAKKA HELA, TRIFI MOKHTAR, OULD MOHAMED SALEM ALI,RHOUMA ABDELMAJID1 and MARRAKCHI MOHAMED∗Laboratoire de Génétique Moléculaire, Immunologie et Biotechnologie, Faculté des Sciencesde Tunis, 2092 El-Manar Tunis, Tunisie; 1Centre de Recherches Phoenicicoles, INRA,Degache, TunisieAbstract. A random genomic library of Tunisian date-palm varieties has been built from total cellular DNA, previously amplified according to an RAPD procedure. The resultant recombinant DNA is characterized by a size ranging from 200 to 1600 bp inserts. This DNA would constitute a large number of anonymous probes useful in Southern hybridization ex- periments. It would also provide potential markers aimed at the molecular characterization of date-palm varieties, aid the search of those associated with bayoud disease and suggest a sex determination of trees. Key words: Date-palm, DNA library, RAPD Abbreviations: BSA, bovine serum albumin; CTAB, hexadecyltrimethylammonium bromide; IPTG, isopropylthio-β-D-galactoside; RAPD, random amplified polymorphic DNA; X-gal, 5- bromo-4-chloro-3-Indolyl-β-D-galactoside. Introduction
Date-palm (Phoenix dactylifera L.), a long-living monocotyledone plant isof economic importance in North Africa. It presents a source of income tooases inhabitants and creates favourable conditions for improving secondarycrop culture like Barley, Alfalfa and Clover as forage. For several decadesdate-palm groves have disappeared almost completely due to a vascular fusar-iosis caused by the fungus, Fusarium oxysporum fsp albedinis (Djerbi, 1982;Djerbi et al., 1985). This fungus has destroyed about 67% of Moroccan palmtrees and has continued to spread to the East, demolishing a large portion ofthe palm groves in its path. Because of the rapid epidemy of bayoud-disease in
∗Author for correspondence. e-mail: [email protected]
the East, Tunisian date-palm plantations are under the threat of complete de-struction. At this time however, Tunisian oases appear to be spared. Even so,the enactment of a preventive defence strategy is keenly encouraged. So far,search of molecular markers associated with the fusariosis have been devel-oped to provide potential early markers (Benslimane et al., 1994; Benslimane,1995).
With the help of RFLP, we have developed a molecular approach to char-
acterize the Tunisian date-palm varieties. This method constitutes a powerfuland reliable technique widely reported to analyse the genetic diversity amonghigher plants (Botstein et al., 1980; Burr et al., 1983; Beckmann and Soller,1986; Tanksley et al., 1987; Mc Couch et al., 1988). Depending on the probeused, various regions of the genome can be targeted in RFLP studies.
Little is known currently about the molecular characterization of date-
palm varieties. Moreover, only Cornicquel and Mercier (1994) have reportedthe use of heterologous probes in Southern hybridization experiments. Be-cause of lacking diversity, it has been assumed that such probes could notprovide a suitable marker. Thus, we have decided to build of a random ge-nomic DNA library from a Tunisian date-palm variety and demonstrate howthis library could provide anonymous markers to be used in RFLP studies.
Here we describe a rapid method aimed at building and screening a ran-
dom genomic library from date-palm. Using this method, we were able toisolate random DNA probes from a genomic DNA library. Material and Methods
• Extraction buffer: 100 mM Tris-HCl, 50 mM EDTA, pH 8.0, 500 mM
• RNase free DNase: 10 mg/ml• Proteinase K: 50 mg/ml• TBE buffer: 89 mM Tris-HCl, 89 mM Boric Acid, 2 mM EDTA, pH 8.0• PCR reaction buffer: 10 mM Tris-HCl, pH 9.0, 50 mM KCl, 1.5 mM
MgCl2, 0.1% Triton X 100, 0.2 mg/ml BSA or Gelatin (Appligène-Oncor, France)
• Ampicillin: 50 mg/ml• X-gal: 20 mg/ml• IPTG: 200 mg/ml
We used the Tunisian date-palm varieties, Tazerzite jaune and Tazerzite noire. They are characterized by an excellent date quality and their abundance inTunisian oases. Plant material, composed of young leaves, was kindly pro-vided by the Centre de Recherches Phoenicicoles, INRA, Degache in theSouth of Tunisia.
Total cellular DNA was extracted according to Dellaporta et al. (1984)
with few modifications. Leaves were ground in liquid nitrogen with a mor-tar and pestle and extracted for 30 min at 65 ◦C in extraction buffer. TheCTAB reagent was used to remove polysaccharides (Lothi et al., 1995) andpolyphenol products (Taylor and Powell, 1985; Ait-Chitt et al., 1993).
For the RAPD procedure, oligonucleotide primers (10 mers) were pur-
chased from Operon Technologies. PCR reactions were performed in a finalvolume of 25 µl and contained 25 ng of total genomic DNA, 10 mM dNTP,50 pM primer and 2 units of Taq DNA polymerase. Amplifications wereperformed in a Thermal Cycler (Bio-med Gm bH) programmed to run 25cycles of [94 ◦C for 30 s, 35 ◦C for 1 min and 72 ◦C for 2 min].
The resultant DNA segments were electroeluted after a preparative gel
electrophoresis. A Sure Clone Ligation Kit (Pharmacia, France) was usedto link this DNA into the Sma I restriction site of pUC 19 according tothe manufacturer’s instructions. After DH5α transformation, bacteria wereplated on LB medium containing 50 mg/ml ampicillin, 20 mg/ml X-gal and200 mg/ml IPTG. White clones were randomly selected from Tazerzite jauneand Tazerzite noire varieties and screened for the presence of recombinantplasmids with inserts greater than 200 bp.
DNA inserts were removed out from pUC 19 vector through restriction
with EcoR I and Hind III enzymes and electrophoresed on 1% agarose gels(Sambrook et al., 1989). Results and Discussion
RAPD assays were performed using OPA07 oligonucleotide as a universalprimer. The amplified products showed an average size of 200 to 1600 bp. These are used in ligation reactions before transformation of DH5α compe-tent cells. A total of 100 of white clones, corresponding to bacteria trans-formed by recombinant plasmids, were selected from each cloning DNAvariety. These plasmid DNAs were also tested for the presence of insertsaccording to their double restriction by EcoR I and Hind III enzymes.
Figure 1 shows typical examples of the resultant recombinant DNA ob-
tained from each variety. It is worth noting that the plasmid DNAs present
Figure 1. Example of restricted recombinant plasmid DNAs deriving from Tazerzite jaune (A)and from Tazerzite noire (B). M: Molecular weight standard size (1 Kb Ladder, Gibco-BRL). Line 1 to 17: Recombinant plasmid DNA restricted by EcoR I / Hind III (for A). Line 1 to 18:Recombinant plasmid DNA restricted by EcoR I / Hind III (for B).
inserts averaging 200 to 1600 bp (for Tazerzite jaune), and of 300 to 1000 bp(for Tazerzite noire). Also, several recombinant clones share inserts charac-terized by similar sizes. A few hypotheses we present to explain these resultsare (1) the inserted DNA is similar in each recombinant plasmid, or (2) theseinserts are different from one given plasmid to another, or (3) both thesehypotheses combined could be correct. Experiments based on the sequencingand/or physical mapping of all these recombinant DNAs would support ourdiscussion.
In addition, Southern (1975) blot hybridization has been performed to
check the date-palm DNA origin inserted in the retained recombinant clones. Thus, using native total cellular DNA as a probe labelled with digoxygenindUTP, hybridization signals are readily detected and correspond to the band-ing patterns from the recombinant DNA (Figure 2).
It is generally believed that genomic clones reveal more polymorphisms
than cDNA clones. The use of cDNA probes has been discussed by Landryet al. (1987), and Miller and Tangsley (1990), with regard to their efficiency indetecting RFLPs in lettuce and tomato. They concluded, however, that cDNAclones reveal more polymorphisms than genomic clones. Figure 2. Hybridization profiles of some recombinant clones deriving from Tazerzite jaunevariety. M: Molecular weight standard size (1 Kb Ladder, Gibco-BRL). Line 1 to 17: Signalsindicating hybridization between inserts and total cellular DNA used as probe.
In Cornicquel and Mercier’s (1994) RFLP analysis of date-palm using
cDNA probes, a polymorphism among cultivars was observed. These cDNAprobes are highly polymorphic and detected polymorphisms in date-palmcultivars.
RFLP technology can be used for the genetic screening of date-palm cul-
tivated groves with economically valuable traits (great yield, fruit quality,resistance to diseases) in date-palm growing countries. Conclusion
Using randomly amplified DNA, we have described a fast, simple and inex-pensive method to produce a reliable genomic DNA library from date-palm. The resultant genomic DNA library, composed of more than fifty recombinantDNAs, corresponds to anonymous probes useful in Southern hybridization. The screening of this library would provide potential markers to molecularlycharacterise date-palm varieties and select those associated in the sex deter-mination of trees and with resistance to the disease. Research is currently inprogress to determine how this recombinant DNA could provide the markerssuitable for studies conducted by any scientist anywhere in the world. Acknowledgements
This work is funded by grants from the Secrétariat d’Etat à la Recherche Sci-entifique et à la Technologie in Tunisia, the Institut Français de la Coopéra-tion (Projet CMCU réf 96/F011) in France and the Tunisian Ministère del’Enseignement Supérieur. References
Ait-Chitt M, Ainsworth CC and Thangavelu M (1993) A rapid and efficient method for ex-
traction of total DNA from mature leaves of date-palm (Phoenix dactylifera L.). Plant MolBiol Reptr 11(4): 317–319.
Beckmann JS and Soller M (1986) Restriction fragment length polymorphism and genetic
improvement of agricultural species. Euphytica 35: 111–124.
Benslimane AA, Rode A, Quetier F and Hartmann C (1994) Characterization of two
minicircular plasmid-like DNAs isolated from date-palm mitochondria. Curr Genet 26:535–541.
Benslimane AA (1995) Plasmides mitochondriaux du palmier dattier (Phoenix dactylifera L.).
Mise en évidence et caractérisation moléculaire. Thèse de Doctorat d’Etat, UniversitéCadi Ayed, Faculté des Sciences Semlalia, Marrakech, Maroc.
Botstein D, White RL, Skolnick M and Davis RW (1980) Construction of genetic linkage map
in man using restriction fragment length polymorphisms. Am J Hum Genet 32: 314–331.
Burr D, Evola SV, Burr F and Beckmann JS (1983) The application of restriction frag-
ment length polymorphism to plant breeding. In: Setlow J, Hollaender A (eds). Geneticengineering. Plenum Press. New York. pp. 45–59.
Cornicquel B and Mercier L (1994) Date-palm (Phoenix dactylifera L.) cultivar identification
by RFLP and RAPD. Plant Science 101: 163–172.
Dellaporta SL, Wood J and Hicks JB (1984) Maize DNA miniprep. In: Molecular Biology of
Plants, Cold Spring Harbor Laboratory Press, New York, pp. 36–38.
Djerbi M (1982) Seminar on bayoud disease in Menadates News 1: 2. Djerbi M, Sedra MH and El Idrisi-Ammari MA (1985) Caractéristiques culturales et identifi-
cation du Fusarium oxysporum fsp albedinis, agent causal du bayoud. Ann Inst Nat RechAgron Tunis 58: 178.
Landry BS, Kesseli R, Leung H and Michelmore RW (1987) Comparison of restriction en-
donucleases and sources of probes for their efficiency in detecting restriction fragmentlength polymorphisms in lettuce (Lactuca sativa L.). Theor Appl Genet 74: 646–653.
Lothi MA, Ye GN, Weeden NF and Reich BI (1994) A simple and efficient method for DNA
extractions from grapevine cultivars and Vitis species. Plant Mol Biol Reptr 12: 6–13.
McCouch SR, Kochert G, Yu ZH, Wang ZY, Khush GS, Coffman WR and Tanksley SD (1988)
Molecular mapping of rice chromosome. Theor Appl Genet 76: 815–829.
Miller JC and Tangsley SD (1990) Effect of different restriction enzymes, probe source, and
probe length on detecting restriction fragment length polymorphism in tomato. Theor ApplGenet 80: 385–389.
Sambrook J, Fritsch EF and Maniatis T (1989) Molecular cloning: a laboratory manual. Cold
Spring Harbor Laboratory Press, New York.
Southern EM (1975) Detection of specific sequences among DNA fragments separated by gel
electrophoresis. J Mol Biol 38: 503.
Taylor B and Powell A (1985) Isolation of plant DNA and RNA. Focus 4: 4–6. Tanksley SD, Miller J, Paterson A and Bernatzky R (1987) Molecular mapping of plant chro-
mosomes. In: Gustafon JP, Appels R (eds) Chromosome structure and function. PlenumPress, New York, pp. 157–173.
Neurohormonal consequences of APS Therapy Study carried out by : Prof. Dr. J.M.C. Oosthuizen MBCHB; DMEDSCI (Head of the Dept. of Physiology; University of the Free Prof. Dr. E.H. de Wet MBCHB; MMED; MD (Dept. of Physiology; University of the Free State) Beta-endorphin, the bodies endogenous analgesic, is a peptide consisting of 31 amino acids with properties similar to morphin
P.O. Box 173460 • Montana State University • Bozeman, MT 59717 (406) 994-2670 • [email protected] ____________________________________________________________________________________________________________ EDUCATION Dissertation: Evaluating alternative approaches to identifying wildlife corridors M.A. Biology: Ecology & Evolutionary Biology University of Califor
 Plant Molecular Biology Reporter 17: 1–7, 2000.
Plant Molecular Biology Reporter 17: 1–7, 2000.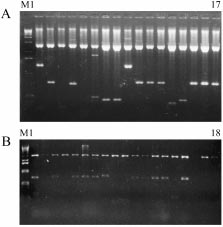 Figure 1. Example of restricted recombinant plasmid DNAs deriving from Tazerzite jaune (A)and from Tazerzite noire (B). M: Molecular weight standard size (1 Kb Ladder, Gibco-BRL).
Figure 1. Example of restricted recombinant plasmid DNAs deriving from Tazerzite jaune (A)and from Tazerzite noire (B). M: Molecular weight standard size (1 Kb Ladder, Gibco-BRL). Figure 2. Hybridization profiles of some recombinant clones deriving from Tazerzite jaunevariety. M: Molecular weight standard size (1 Kb Ladder, Gibco-BRL). Line 1 to 17: Signalsindicating hybridization between inserts and total cellular DNA used as probe.
Figure 2. Hybridization profiles of some recombinant clones deriving from Tazerzite jaunevariety. M: Molecular weight standard size (1 Kb Ladder, Gibco-BRL). Line 1 to 17: Signalsindicating hybridization between inserts and total cellular DNA used as probe.