Die Struktur von Tadalafil erlaubt eine selektive Bindung an die Bindungsstelle der PDE5 und minimiert gleichzeitig die Interaktion mit PDE6, was visuelle Nebenwirkungen einschränkt. Seine Verteilung im Organismus erfolgt breit, wobei das Verteilungsvolumen etwa 63 Liter beträgt. Über 90 % des Wirkstoffs sind an Plasmaproteine gebunden. Die Wirkung bleibt unabhängig von der Nahrungsaufnahme konstant. Der Abbauweg über CYP3A4 kann durch Hemmer wie Ritonavir oder Ketoconazol verlangsamt werden, was die Plasmakonzentrationen deutlich erhöht. In diesem Kontext wird cialis 20mg preis häufig in Bezug auf pharmakokinetische Wechselwirkungen erwähnt.
Olanzapine protects pc12 cells from oxidative stress induced by hydrogen peroxide
Journal of Neuroscience Research 73:364 –368 (2003) Olanzapine Protects PC12 Cells From Oxidative Stress Induced by Hydrogen Peroxide Zelan Wei,1 Ou Bai,1 J. Steven Richardson,1,2 Darrell D. Mousseau,1 and Xin-Min Li1* 1Neuropsychiatric Research Unit, Department of Psychiatry, University of Saskatchewan, Saskatoon, Saskatchewan, Canada 2Department of Pharmacology, College of Medicine, University of Saskatchewan, Saskatoon, Saskatchewan, Canada
Neuroanatomical studies suggest that neuronal atrophy
many other pharmacologic properties that may contribute
and destruction occur over the course of many years in
to their superior therapeutic actions in schizophrenia and
neurodegenerative conditions such as schizophrenia and
that could be the basis for their usefulness in controlling
Alzheimer’s disease. In schizophrenia, early intervention
psychotic symptoms in other disorders as well (Tran et al.,
with atypical neuroleptics such as olanzapine has been
1997; Stoppe and Staedt, 1999; Wolfgang, 1999). Olan-
shown to prevent development of some of the more
zapine, one of the atypical antipsychotics, has been shown
serious and debilitating symptoms in many patients. The
to be safe and effective in reducing behavioral disturbances
mechanisms whereby olanzapine slows or prevents
and psychotic symptoms in patients with Alzheimer’s dis-
symptom progression in schizophrenia remain unclear. A
ease (Street et al., 2000). The mechanisms by which olan-
previous study found that olanzapine increased mRNA
zapine exerts its antipsychotic actions in schizophrenia and
for the copper/zinc isoform of the superoxide dismutase
in Alzheimer’s disease likely involve the blockade of do-
enzyme (SOD-1). We investigated the effects of olanza-
pamine and serotonin receptors; however, the molecular
pine in PC12 cells exposed to hydrogen peroxide. We
mechanisms by which olanzapine and the other atypical
measured cell viability, observed evidence of necrosis
antipsychotics prevent symptom progression remain to be
and apoptosis, checked the SOD-1 mRNA by Northern
blot analyses, and determined SOD-1 enzyme activity.
Neuroanatomical studies suggest that neuronal atro-
We found that: 1) the decrease in cell viability induced by
phy and cell loss occur over many years during the pro-
hydrogen peroxide was attenuated in PC12 cells pre-
gression of neurodegenerative diseases. Neuronal cell
treated with olanzapine; 2) olanzapine increased SOD
death can be induced in many ways. Apoptosis, which is
enzyme activity in PC12 cells; 3) inhibiting SOD activity
involved in normal embryological development of the
with diethyldithiocarbamic acid prevented the cytopro-
central nervous system, also seems to play roles in the
tective actions of olanzapine; and 4) the decrease in
process of normal aging and in the pathophysiology of
SOD-1 mRNA level induced by hydrogen peroxide was
neurodegenerative diseases (Jesberger and Richardson,
blocked by pretreatment with olanzapine. These data
1991; Simonian and Coyle, 1996; Tong and Perez-Polo,
indicate that the neuroprotective action of olanzapine
1996). The apoptotic cell death pathway can be activated
2003 Wiley-Liss, Inc.
by oxidative stress (Jacobson, 1996), and this contributes tothe neurodegenerative process in Alzheimer’s disease
Key words: SOD-1; neuroleptics; hydrogen peroxide;
(Subbarao et al., 1990; Coyle and Puttfarcken, 1993;
Richardson, 1993; Markesbery, 1997). Hydrogen perox-ide, formed as a natural byproduct of enzymatic oxidaseaction, is an endogenous source of free radicals that con-
Although classic antipsychotic drugs such as haloper-
tributes to the background level of cellular oxidative stress
idol produce a marked reduction in positive symptoms ofschizophrenia, they do not improve the negative symp-toms such as apathy, confusion, and social withdrawal, nor
Contract grant sponsor: Canadian Institutes of Health Research (Rx&DResearch Program); Contract grant number: 200104DFE-91579-112515.
do they alter the progressive deterioration in the mentalabilities of the patient. In recent years, several new drugs
*Correspondence to: Dr. X-M Li, Neuropsychiatric Research Unit, 103
have been shown to improve both positive and negative
Wiggins Road, Saskatoon, SK, Canada S7N 5E3.
symptoms of schizophrenia, and seem to prevent further
worsening of psychotic symptoms (Buckley, 1997; Blin,
Received 15 January 2003; Revised 9 April 2003; Accepted 10 April 2003
1999). These atypical antipsychotics are potent antagonistsof both dopamine and serotonin receptors, and also have
2003 Wiley-Liss, Inc. Olanzapine Reduces Oxidative Stress
(Halliwell, 1992; Richardson et al., 1992). Exogenous
from the flask surface with a flow of medium from a pipette, and
hydrogen peroxide can elevate oxidative stress beyond the
dispersing them through a 22-gauge needle. The dispersed
protective capacity of endogenous antioxidant defenses
PC12 cells were then seeded on collagen-coated 96-well plates
and can induce apoptosis and necrosis in cultured cortical
at a cell density of 2 ϫ 104 cells/well, or at the same density (2 ϫ
neurons (Koh et al., 1995; Whittemore et al., 1995) and
105 cells/mL) in T75 tissue flasks. After 24 hr, the cells in the
PC12 cell lines (Tong and Perez-Polo, 1996; Maroto and
96-well plates were treated with olanzapine to give final con-
centrations of 0, 6, 12, 25, 50, 100, or 200 M. After 24 hr of
The rat pheochromocytoma (PC12) cell line is a
exposure to olanzapine, hydrogen peroxide was added to give
well-established model for studying many aspects of the
final concentrations of 0, 100, 200, or 400 M. Other PC12
cellular biology of neurons. PC12 cells have been widely
cells were treated with 100 M olanzapine plus 1 mM diethyl-
used to investigate the mechanisms involved in neurotox-
dithiocarbamic acid for 24 hr, followed by 200 M hydrogen
icity, neuroprotection, and neuronal repair. Oxidative
peroxide. In both cases, after exposure to hydrogen peroxide for
stress induced by hydrogen peroxide (Jackson et al., 1992)
2 hr, cell viability was determined by the MTT assay as de-
or by -amyloid (Zhou et al., 1996) reduces the viability
scribed below. The PC12 cells in T75 flasks were treated with
of PC12 cells, and these effects can be attenuated by nerve
olanzapine (0 or 100 M) for 24 hr and then were exposed to
growth factor or by antioxidant drugs, respectively. Su-
hydrogen peroxide (0 or 100 M). Cells were harvested 1 hr
peroxide dismutase-1 (SOD-1), one of the endogenous
later and examined with the Annexin-V-Fluos Kit. Other cells,
antioxidant enzymes, is also important in protecting cells
exposed to hydrogen peroxide for 2 hr, were harvested and
from oxidative stress. Reducing SOD-1 activity, either by
SOD-1 mRNA levels were determined by Northern blot anal-
downregulating SOD-1 with antisense oligonucleotides or
ysis (see below). SOD activity was determined in PC12 cells
by inhibiting SOD-1 with diethyldithiocarbamic acid, in-
treated with 100 M olanzapine, with 1 mM diethyldithiocar-
duces apoptotic degeneration in neuronal cultures (Roth-
bamic acid, or with both, for 24 hr. In all cases, the olanzapine
stein et al., 1994; Pappolla et al., 1997). Conversely,
and hydrogen peroxide solutions were freshly prepared in
augmenting endogenous SOD-1 activity by administering
RPMI 1640 medium immediately before use.
a metalloporphyrin compound that mimics the action ofSOD protects neurons from staurosporine-induced apo-
Cell Viability Studies
ptosis (Patel, 1998). Previous reports from this laboratory
The viability of PC12 cells in 96-well plates treated as
indicated that olanzapine increases SOD-1 mRNA ex-
above was determined by an MTT assay modified from Mos-
pression in PC12 cells (Li et al., 1999). In the present
mann (1983) and Shearman et al. (1994). In this assay, a yellow
study, we investigate the ability of olanzapine to protect
dye, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-
PC12 cells from oxidative stress induced by hydrogen
mide (MTT), is metabolized by active mitochondria in healthy
cells to a deeply-colored formazan product. Thus, the accumu-lation of formazan is directly proportional to the viability of the
MATERIALS AND METHODS
cells. After the experimental manipulations outlined above, theold medium was removed from each well and replaced with 50
Materials
L of RPMI-1640 with 1% newborn calf serum containing 0.5
The PC 12 cell line, obtained from American Type Cul-
mg/mL MTT in 0.1 M phosphate-buffered saline (PBS). The
ture Collection (Rockville, MD), was grown in RPMI 1640
96-well plates were incubated at 37°C for 4 hr, and then 100 L
culture medium obtained from local sources. Newborn calf
of acid-isopropanol (0.04 M HCl) was added to each well. After
serum, horse serum, diethyldithiocarbamic acid and all other
an additional 4 hr at 37°C to insure that all formazan crystals had
chemicals were purchased from Sigma Chemical Co. (St. Louis,
been dissolved, the optical density of each well was determined
MO). The Annexin-V-Fluos Staining Kit for detecting apopto-
with a microELISA reader at a wavelength of 570 nm. In this
sis was purchased from Roche Molecular Biochemicals (Laval,
assay, cell viability is proportional to the optical density of each
PQ, Canada). TRIZOL reagent was purchased from Life Tech-
well. After exposure to olanzapine (0 or 100 M) for 24 hr, and
nologies (Rockville, MD). The SOD Assay Kit-WST was pur-
hydrogen peroxide (0 or 100 M) for 1 hr, cells were examined
chased from Dojindo Molecular Technologies (Kumamoto, Ja-
with the Annexin-V-Fluos Staining Kit, a histological procedure
pan), and the BCA Protein Assay Reagent kit for measuring
that differentially labels necrotic cells versus cells undergoing
protein concentration was from Pierce Inc. (Rockford, IL).
apoptosis. Although the MTT assays were done after 2 hr of
Olanzapine was a gift from Lilly Research Laboratories (India-
exposure to H O , preliminary studies showed that 1 hr of
exposure to H O produced optimal resolution with Annexin-
Cell Culture and Experimental Treatment
The PC12 cells were cultured in RPMI 1640 medium
SOD Studies
containing 5% newborn calf serum and 10% horse serum plus
Northern Blot.
100 IU/mL penicillin G sodium salt and 3% glutamine. Cells
Dr. J.T. Coyle (Harvard Medical School, Boston, MA). The
were fed with fresh medium every 2 or 3 days and maintained
cDNA probes were labeled by random primer synthesis with
in collagen-coated T75 tissue culture flasks at 37°C in a humid-
[␣-32P] dCTP as described previously (Li et al., 1999). Briefly,
ified atmosphere containing 5% CO and 95% air. When cells
total cellular RNA was prepared from treated cells by extraction
became 80% confluent, they were harvested by dislodging them
with TRIZOL reagent according to supplier’s instructions. The
Wei et al.
RNA was resuspended in DEPC-treated water and stored at
Ϫ70°C until use. The total RNA, denatured at 65°C for 15 minin MOPS buffer containing 50% formamide and 2.2 M form-aldehyde, was separated by electrophoresis in a 1.0% agarose gelcontaining MOPS buffer and 2.2 M formaldehyde. After elec-trophoresis, RNA was transferred to a nylon membrane andcross-linked to the membrane with a UV Stratalinker 2400.
The nylon membranes were incubated at 65°C for 2 hr in
a prehybridization solution containing 10% dextran sulfate, 5ϫSSPE, 5ϫ Denhardt’s solution, 0.5% SDS, and denaturedsalmon sperm DNA (200 g/mL). The cDNA probe was addedand hybridization was carried out at 65°C for 18 hr. Afterhybridization, membranes were washed twice at room temper-ature in 2ϫ SSPE and 0.1% SDS, and once at 60°C in 0.1ϫSSPE and 0.1% SDS. The membranes were then exposed atroom temperature to X-Omat AR film with intensifying screens
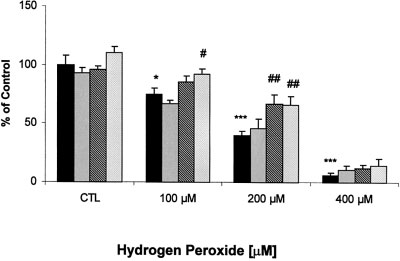
Fig. 1. Effects of olanzapine on H O -induced neurotoxicity in PC12
to obtain autoradiograms of the membranes. The autoradio-
cells. PC12 cells were treated with olanzapine for 24 hr in different
grams were scanned with a computerized densitometer (Du 640,
concentrations, 0 (solid black), 50 (cross-hatched), 100 (diagonal), or
Beckman Inc., Fullerton, CA) for quantitative analysis and the
200 (blank) M, then exposed to 0, 100, 200, or 400 M H O for
signals were adjusted according to the original signal in trans-
2 hr. Cell viability was measured with the MTT assay. Data (mean Ϯ
SEM of five separate experiments) are expressed as a percentage of the
SOD Enzyme Activity
control value. *P Ͻ 0.05, ***P Ͻ 0.001, compared to the untreated
harvested and lysed in a hypotonic buffer (1% NP-40, 50 mM
control value. #P Ͻ 0.05, ##P Ͻ 0.01, compared to correspondingH O concentrations without olanzapine.
Tris-HCl, pH 7.5, 0.05 mM EDTA) for 20 minutes at 4°C. The
lysates were centrifuged at 16,000 ϫ g for 10 minutes, and analiquot of the supernatant was used for determination of SODactivity with the SOD Assay Kit-WST following the manufac-turer’s instructions (Shimazaki et al., 2002). Protein concentra-tions of cell lysates were measured using the BCA (bicinchoninicacid) Protein Assay Reagent Kit. The SOD activity was adjustedby the amount of protein in each sample. Statistical Analyses
All data points are presented as means Ϯ SEM. Statistical
significance was assessed by ANOVA followed by Newman-Keul’s post-hoc comparisons using Prism software (GraphPadSoftware Inc., San Diego, CA).
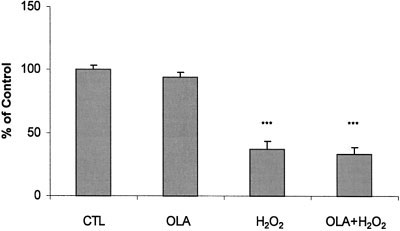
Fig. 2. Effects of diethyldithiocarbamic acid on the action of olanzap-
Effect of Olanzapine on Viability of PC12 Cells
ine and H O on PC12 cells. Cells were treated with 1 mM diethyl-
Exposed to H O
dithiocarbamic acid plus 0 or 100 M olanzapine for 24 hr, and then
The ANOVA indicated that the main effects of
exposed to 0 or 200 M H O for 2 hr. Cell viability was measured
hydrogen peroxide (F ϭ 187; df ϭ 3, 48; P Ͻ 0.001) and
with the MTT assay. Data (mean Ϯ SEM of four separate experiments)
of olanzapine (F ϭ 8.4; df ϭ 3, 48; P Ͻ 0.001) on the
are expressed as a percentage of the diethyldithiocarbamic acid alone
viability of PC12 cells were statistically significant. As
control value. ***P Ͻ 0.001, compared to the control value.
shown in Figure 1, the exposure of PC12 cells to increas-ing concentrations of H O
1 mM of the SOD inhibitor diethyldithiocarbamic acid,
dependent decrease in cell viability, whereas olanzapine, at
100 M olanzapine did not attenuate the 60% reduction
the concentrations used, did not alter cell viability. The
in viability induced by 200 M hydrogen peroxide (Fig.
viability of PC12 cells exposed to 100 M hydrogen
2). Annexin-V-Fluos staining indicated that numerous
peroxide was reduced by 25%. This was not altered by
PC12 cells were undergoing apoptosis or necrosis after
50 M olanzapine, but was prevented by the 100 and
treatment with H O , whereas there were fewer apoptotic
200 M concentrations of olanzapine. The 60% reduction
cells in olanzapine pretreated cultures (data not shown).
in viability produced by 200 M hydrogen peroxide was
attenuated by the two higher concentrations of olanzap-
Investigation of SOD-1 in PC12 Cells Treated
ine. None of the olanzapine concentrations used was able
With Olanzapine and H O
to alter the 90% reduction in viability produced by
As shown in Figure 3, the expression of SOD-1
400 M hydrogen peroxide. In PC 12 cells treated with
mRNA was reduced by 30% by 100 M hydrogen per-
Olanzapine Reduces Oxidative Stress
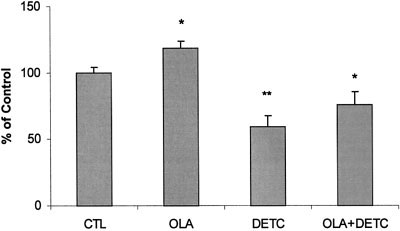
Fig. 4. Effects of diethyldithiocarbamic acid (DETC) and olanzapineon SOD-1 activity in PC12 cells. Cells were treated with vehicle alone,100 M olanzapine, 1 mM diethyldithiocarbamic acid, or both, for24 hr. SOD-1 activity was measured according to the SOD AssayKit-WST. Data (mean Ϯ SEM of three to five separate experiments)are expressed as a percentage of the vehicle control value. *P Ͻ 0.05,**P Ͻ 0.01 compared to control value.
Fig. 3. Effect of olanzapine on SOD-1 mRNA expression in PC12
sumption that predisposes to free radical formation, and,
cells. Cells were pretreated with olanzapine (0 or 100 M) for 24 hr,
compared to other organs, is relatively deficient in anti-
and then exposed for 2 hr to 0 or 100 M H O . A: The Northern blot
oxidant activity (Floyd, 1999). When the production of
bands show SOD-1 mRNA expression in total cell lysates. B: Data,
free radicals exceeds the protective capacity of the cellular
expressed as percentage of untreated control, are the means Ϯ SEM of
antioxidant defenses, this leads to oxidative damage to
the integrated density volumes of four separate Northern blot experi-
neuronal proteins and lipids, which reduces the efficiency
ments. *P Ͻ 0.05, compared to the untreated control, # P Ͻ 0.05,
of neuronal communication, and, if severe enough, will
induce neuronal apoptosis. Cu/Zn superoxide dismutase(SOD-1) is one of the key enzymes by which cells detox-ify free radicals and protect themselves from oxidative
oxide. If cells were pretreated with 100 M olanzapine
before exposure to hydrogen peroxide, however, SOD-1
Because olanzapine increases SOD activity (Fig. 4),
mRNA expression was not significantly decreased.
the neuroprotective effects of olanzapine (Fig. 1) may be
SOD-1 enzyme activity (Fig. 4) was increased by 10%
due to acceleration of metabolic processing of free radicals.
after treatment with 100 M olanzapine for 24 hr, and was
As indicated by Northern blot studies, olanzapine not only
reduced by 40% after 24 hr of exposure to 1 mM dieth-
increases SOD-1 mRNA (Li et al., 1999), but also inhibits
yldithiocarbamic acid. Olanzapine did not significantly
the decrease in SOD-1 mRNA caused by H O (Fig. 3).
alter the SOD inhibition produced by diethyldithiocar-
When SOD-1 activity is inhibited by diethyldithiocar-
bamic acid (Fig. 4), the protective effect of olanzapine islost. This indicates that elevated SOD-1 activity plays a
DISCUSSION
role in the cytoprotective action of olanzapine; however,
Although it is known that clinically efficacious anti-
the underlying mechanisms by which olanzapine increases
psychotics block central dopamine D receptors, seroto-
SOD-1 activity and how this reduces cell damage remain
nergic 5-HT receptors, or both, the total clinical effect of
to be determined. Because the annexin staining study
these drugs may depend on other actions in addition to
revealed less evidence of apoptosis, but no change in
receptor blockade. The present study examines the effect
necrosis, in cultures pretreated with olanzapine before
of olanzapine on the regulation of SOD activity in PC12
exposure to hydrogen peroxide, it seems that olanzapine
cells exposed to excessive levels of oxygen free radicals.
can protect cells from apoptosis, but not necrosis.
Oxidative stress contributes to neuronal destruction
Future work will focus on how olanzapine regulates
in a wide range of neurodegenerative disorders, such as
posttranslational modification of SOD-1 and will investi-
cerebral ischemia, Alzheimer’s disease, Parkinson’s disease,
gate the effects of olanzapine on other aspects of the
and amyotrophic lateral sclerosis (Jesberger and Richard-
cellular oxidative stress protection system.
son, 1991; Behl, 1997; Markesbery, 1997). A variety offactors make brain function especially vulnerable to dis-
ACKNOWLEDGMENTS
ruption by oxidative stress. The brain has a high metabolic
We thank Ms. G. Stegeman for her technical assis-
rate with a correspondingly high level of oxygen con-
tance. This work was supported in part by a postdoctoral
Wei et al.
fellowship from the CIHR/Rx&D Research Program to
Richardson JS. 1993. Free radicals in the genesis of Alzheimer’s disease.
Richardson JS, Subbarao KV, Ang LC. 1992. On the possible role of
REFERENCES
iron-induced free radical peroxidation in neural degeneration in Alzhei-
Behl C. 1997. Amyloid -protein toxicity and oxidative stress in Alzhei-
mer’s disease. Ann N Y Acad Sci 648:326 –327.
mer’s disease. Cell Tissue Res 290:471– 480.
Rothstein JD, Bristal LA, Hosler B, Brown RH Jr, Kuncl RW. 1994.
Blin O. 1999. A comparative review of new antipsychotics. Can J Psychi-
Chronic inhibition of superoxide dismutase produces apoptotic death of
spinal neurons. Proc Natl Acad Sci USA 91:4155– 4159.
Buckley PF. 1997. New dimensions in the pharmacologic treatment of
Shearman MS, Ragan CI, Iversen LL. 1994. Inhibition of PC12 cell redox
schizophrenia and related psychoses. J Clin Pharmacol 37:363–378.
activity is a specific, early indicator of the mechanism of -amyloid-
Coyle JT, Puttfarcken P. 1993. Oxidative stress, glutamate, and neurode-
mediated cell death. Proc Natl Acad Sci USA 91:1470 –1474.
generative disorders. Science 262:689 – 695.
Shimazaki Y, Ohnishi H, Matsuura S, Manabe T. 2002. Simple quantifi-
Floyd RA. 1999. Antioxidants, oxidative stress, and degenerative neuro-
cation of Cu, Zn-superoxide dismutase activity after separation by non-
logical disorders. Proc Soc Exp Biol Med 222:236 –245.
denaturing isoelectric focusing. Biochim Biophys Acta 1571:245–248.
Halliwell B. 1992. Reactive oxygen species and the central nervous system.
Simonian NA, Coyle JT. 1996. Oxidative stress in neurodegenerative
diseases. Ann Rev Pharmacol Toxicol 36:83–106.
Jackson GR, Werrbach-Perez K, Ezell EL, Post JF, Perez-Polo JR. 1992.
Stoppe G, Staedt J. 1999. [Psychopharmacotherapy of behavioral disorders
Nerve growth factor effects on pyridine nucleotides after oxidant injury of
in patients with dementia.] German. Z Gerontol Geriatr 32:153–158.
rat pheochromocytoma cells. Brain Res 592:239 –248.
Street J, Clark WS, Gannon KS, Cummings JL, Bymaster FP, Tamura RN,
Jacobson MD. 1996. Reactive oxygen species and programmed cell death.
Mitan SJ, Kadam DL, Sanger TM, Feldman PD, Tollefson GD, Breier A.
2000. Olanzapine treatment of psychotic and behavioral symptoms in
Jesberger JA, Richardson JS. 1991. Oxygen free radicals and brain dysfunc-
patients with Alzheimer disease in nursing care facilities: a double-blind,
randomized, placebo-controlled trial. The HGEU Study Group. Arch
Koh JY, Gwag BJ, Lobner D, Choi DW. 1995. Potentiated necrosis of
cultured cortical neurons by neurotrophins. Science 268:573–575.
Subbarao KV, Richardson JS, Ang LC. 1990. Autopsy samples of Alzhei-
Li XM, Chlan-Fourney J, Juorio AV, Bennett VL, Shrikhande S, Keegan
mer’s cortex show increased peroxidation in vitro. J Neurochem 55:342–
DL, Qi J, Boulton AA. 1999. Differential effects of olanzapine on the
gene expression of superoxide dismutase and the low affinity nerve
Tong L, Perez-Polo JR. 1996. Effect of nerve growth factor on AP-1,
growth factor receptor. J Neurosci Res 56:72–75.
NF-B, and Oct DNA binding activity in apoptotic PC12 cells: extrinsic
Markesbery WR. 1997. Oxidative stress hypothesis in Alzheimer’s disease.
and intrinsic elements. J Neurosci Res 45:1–12.
Tran PV, Hamilton SH, Kuntz AJ, Potvin JH, Andersen SW, Beasley C Jr,
Maroto R, Perez-Polo JR. 1997. BCL-2-related protein expression in
Tollefson GD. 1997. Double-blind comparison of olanzapine versus
apoptosis: oxidative stress versus serum deprivation in PC12 cells. J Neu-
risperidone in the treatment of schizophrenia and other psychotic disor-
ders. J Clin Psychopharmacol 17:407– 418.
Mosmann T. 1983. Rapid colorimetric assay for cellular growth and sur-
Whittemore ER, Loo DT, Watt JA, Cotman CW. 1995. A detailed analysis
vival: application to proliferation and cytotoxicity assays. J Immunol
of hydrogen peroxide-induced cell death in primary neuronal culture.
Pappolla MA, Sos M, Omar RA, Bick RJ, Hickson-Bick DL, Reiter RJ,
Wolfgang SA. 1999. Olanzapine in whole, not half, tablets for psychosis
Efthimiopoulos S, Robakis NK. 1997. Melatonin prevents death of
from Alzheimer’s dementia. Am J Health Syst Pharm 56:2245–2246.
neuroblastoma cells exposed to the Alzheimer amyloid peptide. J Neu-
Zhou Y, Gopalakrishnan V, Richardson JS. 1996. Actions of neurotoxic
-amyloid on calcium homeostasis and viability of PC12 cells are blocked
Patel M. 1998. Inhibition of neuronal apoptosis by a metalloporphyrin
by antioxidants but not by calcium channel antagonists. J Neurochem
superoxide dismutase mimic. J Neurochem 71:1068 –1074.
Extraction of Caffeine from Tea Purpose is to learn some of the basic techniques of organic chemistry: extraction, filtration, evaporation of a solvent and drying methods-in the context of working with a chemical known to all, caffeine. Pure caffeine is a white, tasteless substance that makes up as much as 5% of the weight of tea leaves. By structure, caffeine is closely related to the
Behandeling spasticiteit Wat is spasticiteit? Spasticiteit is een snelheidsafhankelijke bewegingsstoornis van spieren, die een te hoge spanning hebben. Spasticiteit wordt veroorzaakt door beschadiging van de hersenen of het ruggenmerg, die het gevolg is van een letsel (bijvoorbeeld ongeval), een aangeboren afwijking of beschadiging bij de geboorte, maar ook door ziekte op latere leeftijd (
 Wei et al.
Wei et al.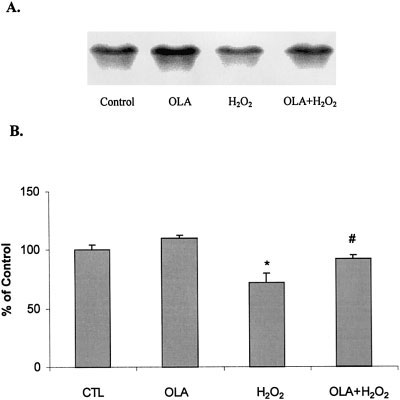
 Olanzapine Reduces Oxidative Stress
Olanzapine Reduces Oxidative Stress