Die Struktur von Tadalafil erlaubt eine selektive Bindung an die Bindungsstelle der PDE5 und minimiert gleichzeitig die Interaktion mit PDE6, was visuelle Nebenwirkungen einschränkt. Seine Verteilung im Organismus erfolgt breit, wobei das Verteilungsvolumen etwa 63 Liter beträgt. Über 90 % des Wirkstoffs sind an Plasmaproteine gebunden. Die Wirkung bleibt unabhängig von der Nahrungsaufnahme konstant. Der Abbauweg über CYP3A4 kann durch Hemmer wie Ritonavir oder Ketoconazol verlangsamt werden, was die Plasmakonzentrationen deutlich erhöht. In diesem Kontext wird cialis 20mg preis häufig in Bezug auf pharmakokinetische Wechselwirkungen erwähnt.
Iah.iec.pa.gov.br
JOURNAL OF CLINICAL MICROBIOLOGY, Aug. 2006, p. 2767–2772
0095-1137/06/$08.00ϩ0 doi:10.1128/JCM.01916-05Copyright 2006, American Society for Microbiology. All Rights Reserved.
Clonality and Antimicrobial Resistance Gene Profiles of Multidrug-
Resistant Salmonella enterica Serovar Infantis Isolates from
Four Public Hospitals in Rio de Janeiro, Brazil
E. L. Fonseca,1 O. L. Mykytczuk,4,5 M. D. Asensi,1 E. M. F. Reis,1 L. R. Ferraz,2
F. L. Paula,3 L. K. Ng,4,5 and D. P. Rodrigues1*
Bacteriology Department, Oswaldo Cruz Institute – FIOCRUZ, Rio de Janeiro, Brazil1; Public Health Laboratory, Brasilia, Brazil2;Evandro Chagas Institute, Para, Brazil3; National Microbiology Laboratory, Public Health Agency of Canada,Winnipeg, Manitoba, Canada4; and Department of Medical Microbiology, Faculty of Medicine,University of Manitoba, Winnipeg, Manitoba, Canada5
Received 13 September 2005/Returned for modification 7 November 2005/Accepted 8 May 2006
In Brazil, Salmonella enterica serovar Infantis resistant to various antimicrobials, including cephalosporins, has been identified as an etiological agent of severe gastroenteritis in hospitalized children since 1994. In this study, 35 serovar Infantis strains, isolated from children admitted to four different Rio de Janeiro, Brazil, hospitals between 1996 and 2001, were characterized by pulsed-field gel electrophoresis (PFGE) and antimi- crobial susceptibility testing in order to determine their genetic relatedness and antimicrobial resistance profiles. Thirty-four serovar Infantis strains were resistant to at least two antibiotic classes, and all 35 strains were susceptible to fluoroquinolones, cephamycin, and carbapenem. Extended-spectrum beta-lactamase (ESBL) screening by double-disk diffusion indicated that 32 serovar Infantis strains (91.4%) produced beta- lactamases that were inhibited by clavulanic acid. Antimicrobial resistance gene profiles were determined by PCR for a subset of 11 multidrug-resistant serovar Infantis strains, and putative ESBLs were detected by isoelectric focusing. Ten serovar Infantis strains carried bla , catI, ant(3؆)Ia and/or ant(3؆)Ib, sulI and/or sulII, and tet(D) genes as well as an integron-associated aac(6)-Iq cassette. Eight strains possessed at least four different beta-lactamases with pI profiles that confirmed the presence of both ESBLs and non-ESBLs. Our PFGE profiles indicated that 33 serovar Infantis strains isolated from Rio de Janeiro hospitals came from the same genetic lineage.
For many years, ampicillin, sulfamethoxazole-trimethoprim,
isolated in three other public health hospitals (designated HA,
and chloramphenicol were the drugs of choice for the treat-
HB, and HD) of Rio de Janeiro, Brazil. Two are pediatric
ment of severe Salmonella infections, but increasing rates of
reference hospitals that often see children from the western
resistance to these agents have significantly reduced their ef-
and northern regions of the city, where parts of the population
ficacies (28, 35). Subsequently, third-generation cephalospo-
have lower socioeconomic and sanitary conditions. Some chil-
rins, due to their pharmacodynamic properties as well as low
dren were human immunodeficiency virus positive, and most
resistance levels in Salmonella, are being used to treat invasive
suffered from recurring infections and had histories of rehos-
pitalization. Although HC is a university-affiliated hospital and
In 1994, Asensi and Hofer reported the presence in Rio de
HD is a reference hospital for cancer, both provide medical
Janeiro, Brazil, of Salmonella enterica serovar Infantis strains
care for patients with debilitating diseases such as AIDS and
that were resistant to a growing number of antimicrobial
diabetes. In addition, these patients are subjected to prolonged
agents (6). Two years later, a nosocomial outbreak in a neo-
hospitalizations that are often accompanied by the empirical
natal unit of one hospital (designated HC) was reported by De
use and sometimes overuse of antimicrobial drugs (ampicillin
Moraes et al. (13). The authors detected multidrug-resistant
and/or cephalosporins and/or aminoglycosides). This led us to
serovar Infantis phenotypes, including resistance to broad-
monitor the prevalence and antimicrobial susceptibility of se-
spectrum cephalosporins that was transferred by a plasmid of
rovar Infantis in hospitals in Rio de Janeiro, Brazil. The aims
148 kbp. An investigation carried out from 1998 to 1999 re-
of this research were to (i) determine the antimicrobial sus-
ported an infection due to extended-spectrum beta-lactamase
ceptibility patterns, (ii) identify the main mechanisms involved
(ESBL)-producing serovar Infantis in the neonatal unit of a
in antimicrobial resistance, (iii) ascertain the presence and
public hospital (HC) in Rio de Janeiro, Brazil, indicating in-
spread of integron-carried resistance genes, and finally, (iv)
adequate infection control practices and nursery overcrowding
assess the macro-restriction fragment length polymorphisms
(30). Since then, multidrug-resistant serovar Infantis has been
between multidrug-resistant serovar Infantis strains from thosehospitals.
* Corresponding author. Mailing address: Laborato
MATERIALS AND METHODS
´rias, Departamento de Bacteriologia, Oswaldo Cruz Institute –
FIOCRUZ, Avenida Brasil, 4365 – Pavilha
Bacterial strains. Serovar Infantis strains were isolated according to the
Manguinhos – Rio de Janeiro, Brasil 21040-361. Phone: 55 21 2598
method of Costa and Hofer (12), and the antigenic characterization was based on
4277. Fax: 55 21 2270 6565. E-mail: [email protected].
the Kauffmann-White scheme described by Poppof (29a). This study included 35
TABLE 1. PCR primers used to identify antimicrobial resistance genes and integrons in serovar Infantis
F, GCT ACA TCC TGC TTG CCT TC; R, CAT AGA
F, TTG GTT AGG GGC AAG TTT TG; R, GTA ATG
F, CTT GAG AGC CTT CAA CCC AG; R, ATG GTC
F, AAA CCA TTA CGG CAT TCT GC; R, GAC CGG
F, AAA CCA CAT CCT CCA TAC GC; R, AAA TAG
F, CAG CTT TCG GAT TCT TAC GG; R, GAT TGG
F, CCT GAA AAC CAA ACT GCC TC; R, ACA GAC
F, TCA GCT GGA TAT TAC GGC CT; R, CAT TCT
F, ATT CAG CCT GAC CAC CAA AC; R, CTT CCT
F, CCC ACA ATT CAC CGT ATT CC; R, GAA CCT
F, CAC CGC GGC GAT CGA AAT GC; R, GGT TTC
F, ATC GCT CAT CAT TTT CGG CA; R, CTC GTG
F, CGA AGA ATG GAG TTA TCG GG; R, TAA ACA
F, GCG CTA AAT GAA ACC TTA AC; R, TCG CCT
F, TGT TGG TTA CTG TGG CCG TA; R, GCT GCG
F, TTA TGC CTC TTC CGA CCA TC; R, GAG AAA
F, GCT GGA AAT GAA TCA TGG GT; R, TAA TTC
F, ATA AAA TTC TTG AAG ACG AAA; R, GAC AGT
F, GGC ATC CAA GCA GCA AG; R, AAG CAG ACT
a F, forward primer; R, reverse primer. b The reference strain served as a positive control for PCRs.
serovar Infantis strains isolated from the stools or blood of children under 7
confirmed at the National Microbiology Laboratory, Public Health Agency of
months who were admitted to four public hospitals (HA, HB, HC, and HD) in
Canada, by using the Mast Diagnostics ESBL detection kit (Merseyside, United
Rio de Janeiro, Brazil, from 1996 to 2001. Only one isolate per patient was
Kingdom) according to the manufacturer’s instructions. Preparation of crude protein extracts and IEF. The 11 ESBL-positive isolates Antimicrobial susceptibility testing and extended-spectrum beta-lactamase
were grown in 2 ml of Mueller-Hinton broth at 37°C overnight, and cells were
assay. Disk diffusion tests were performed according to Clinical and Laboratory
harvested by centrifugation at 16,000 ϫ g for 2 min. After discarding the super-
Standards Institute (formerly National Committee for Clinical Laboratory Stan-
natant, cells were resuspended in 250 l of 1% glycine and 30% glycerol and
dards) (26) recommendations by using disks (Oxoid Limited, Hampshire, England)
were sonicated twice for 30 s, with cooling of the cells on ice between sonications.
impregnated with ampicillin (AMP; 10 g), aztreonam (ATM; 30 g), cephalothin
Cell lysates were centrifuged at 16,000 ϫ g for 15 min. Supernatants were
(CEF; 30 g), cefotaxime (CTX; 30 g), ceftriaxone (CRO; 30 g), ceftazidime
collected into clean tubes and stored at Ϫ20°C. Prior to isoelectric focusing
(CAZ; 30 g), cefoxitin (FOX; 30 g), cefuroxime (CXM; 30 g), cefepime (FEP;
(IEF), cell extracts were tested for beta-lactamase activity by adding 50 l of 50
30 g), ciprofloxacin (CIP; 5 g), chloramphenicol (CHL; 30 g), streptomycin
g/ml nitrocefin stock solution (Oxoid Limited, England) to 17 l of extract and
(STR; 10 g), kanamycin (KAN; 10 g), gentamicin (GEN; 10 g), imipenem (IPM;
then recording the time required for the reaction to turn dark pink. The optimal
10 g), nalidixic acid (NAL; 30 g), trimethoprim-sulfamethoxazole (SXT; 25 g),
reaction time was 30 to 120 s. For reaction times of 5 s or less, the extract was
and tetracycline (TET; 30 g). For quality control of the culture media and antimi-
diluted with phosphate buffer and retested. For isolates with reaction times of 5
crobial disks, Escherichia coli ATCC 25922, E. coli ATCC 35218, Pseudomonas
min or more, another extract was prepared from a culture of greater density and
aeruginosa ATCC 27853, Enterococcus faecalis ATCC 29212, and Staphylococcus
the test was redone. For IEF, precast polyacrylamide IEF minigels (pH 3 to 10)
aureus ATCC 25923 were tested under the same conditions and antimicrobials as
(Bio-Rad Laboratories, Hercules, CA) were assembled in a vertical Bio-Rad
Mini-Protean II electrophoresis unit. Cathode buffer (20 mM lysine-20 mM
The method described by the CLSI for “other Enterobacteriaceae” was used to
arginine) (Bio-Rad) was added to the middle chamber, the wells were flushed,
perform double-disk diffusion for the screening of ESBL-producing strains. Dou-
and then 10 l of crude extract was loaded in every second well. An IEF standard
ble-disk diffusion was performed with cephalosporin and cephalosporin/clavu-
with pIs ranging from 4.45 to 9.6 (Bio-Rad) was used, and a marker composed
lanic acid combination disks (Oxoid Limited, England). Klebsiella pneumoniae
of beta-lactamases of known isoelectric points (pIs) (blaTEM-1 [pI 5.4], blaTEM-4
ATCC 700603 (positive) and E. coli ATCC 25922 (negative) were used as control
[pI 5.9], blaTEM-3 [pI 6.3], blaSHV-3 [pI 7.0], and blaSHV-2 [pI 7.6]) was also used.
strains. In addition, the production of ESBLs in 11 serovar Infantis strains was
Approximately 200 ml of anode buffer (7 mM phosphoric acid) (Bio-Rad) was
GENE PROFILES OF SALMONELLA ENTERICA SEROVAR INFANTIS
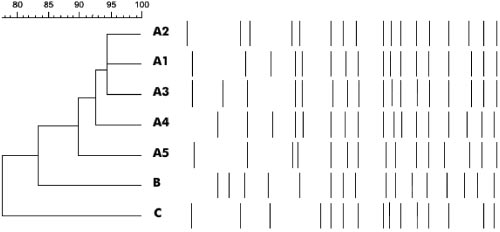
FIG. 2. PFGE macro-restriction fragment polymorphism.
gel run at 100 V for 1 h. To visualize band migration, the gel was stained withethidium bromide and observed under UV light. A 100-bp or 1-kb ladder (GibcoBRL, Ontario) was used to estimate amplicon size. DNA sequencing. Amplicons resulting from PCRs using the primers specific to
the 5Ј conserved and 3Ј semiconserved segments or universal blaTEM primerswere sequenced in both directions using an ABI Prism 377 DNA sequencer(Applied Biosystems Division of Perkin-Elmer, Foster City, CA). DNA se-quences were compared to those in the GenBank database (National Center forBiotechnology Information) by using the BLAST suite of sequence similarity-searching programs (3, 4). Pulsed-field gel electrophoresis (PFGE). Genomic DNA was prepared as
described previously by Persing et al. (29) with modifications. Serovar Infantisstrains were grown in 10 ml of Mueller-Hinton broth at 37°C for 12 to 18 h. Cellswere harvested by centrifugation at 2,000 ϫ g for 15 min. After discarding thesupernatant, cells were resuspended with 1 ml of sterilized saline (0.85% NaCl)
FIG. 1. PFGE: macro restriction fragment patterns of Salmonella
and the concentration was adjusted to 1 ϫ 106 cells/ml. A 5-l aliquot of cell
serovar Infantis genome digested with SpeI. Lanes: M, molecular
suspension was added to 300 l of TEN buffer (0.5 M EDTA, 1 M Tris base, 4
weight marker of Salmonella Branderup strains; A2, PFGE profile of
M NaCl, pH 7.5) before embedding it in 340 l of low-melting-point agarose
3 HD strains; C, PFGE profile of a midwestern hospital strain; B,
(Sigma-Aldrich Corporation, St. Louis, MS). Plugs were subjected to lysis for 5 h
PFGE profile of a northern hospital strain; A1, PFGE profile of 18
at 37°C in EC buffer (0.5 M EDTA, 1 M Tris base, NaCl, N-lauryl sarcosyl, Brij
HA, 4 HB, 5 HC strains; A3, PFGE profile of 1 HA strain; A5, PFGE
58, sodium deoxycholate, pH 7.0) (Sigma-Aldrich, MS). RNase (10 mg/ml)
profile of 1 HA strain; A4, PFGE profile of 1 HA strain.
(Sigma-Aldrich, MS) was added to the plugs for an overnight incubation at 37°C,and then proteinase K (20 mg/ml; Gibco BRL) treatment of the plugs wasperformed for 24 h at 54°C. Serovar Infantis strain plugs were washed four timeswith CHEF-TE 1ϫ buffer (0.5 M EDTA, 1 M Tris base, pH 7.5) (Sigma-Aldrich,
added to the outer buffer chamber. The electrophoresis unit was placed on a tray
MS), followed by four washes with DNS buffer (1 M Tris base, 1 M MgCl2)
and surrounded with ice. Electrophoresis was performed in three steps: 100 V for
(Sigma-Aldrich, MS). The digestion step was performed for 20 h at 37°C with the
1 h, 250 V for another hour, and finally, 500 V for 30 min. IEF gels were then
restriction endonuclease SpeI (10 U/l) (Amersham Pharmacia Biotech, En-
dismantled from the unit, and the glass plates were separated while leaving the
gland). Electrophoresis was performed at 6 V/cm for 22 h with switch time
gel on one glass plate. To visualize beta-lactamase activity, 1 ml of nitrocefin
intervals of 0.5 to 25 s for 19 h and 30 to 60 s for 3 h on CHEF DRIII (Bio-Rad
stock solution (1 mg/ml) was added to 6 ml of molten 3% agarose in 50 mM
Laboratories, Richmond, CA). The agarose gels were stained with ethidium
phosphate buffer (pH 7.5) (cooled to 50 to 60°C), mixed by inversion, and then
bromide, visualized by UV transillumination, and photographed on Image-
poured evenly over the gel. The presence of pink/red lines on the gel indicated
Master VDS (Amersham Pharmacia Biotech, England). The fragment restriction
beta-lactamase activity. Pictures of IEF gels were taken using a dark green filter,
patterns were analyzed by BioNumerics (Applied Maths, Belgium) and com-
and the gels were transilluminated with white light.
pared through the construction of a similarity matrix by using the Dice coefficient
Detection of antimicrobial resistance genes. PCR was used to detect antimi-
with a position tolerance setting of 1.0% and optimization setting of 1.0%, which
crobial resistance genes and the presence of integrons in 11 isolates resistant to
generated a dendrogram. Serovar Branderup was included as a control. A clonal
(at least) the following antimicrobials: ampicillin, chloramphenicol, streptomy-
structure definition of serovar Infantis was achieved according to the criteria of
cin, sulfamethoxazole-trimethoprim, and tetracycline. Most of the primers used
Tenover et al., which correlates the number of fragment differences with genetic
for the characterization of pentaresistant Salmonella serovar Typhimurium
DT104 were previously described (27), and are all listed in Table 1. The DNA
Two human epidemiologically unrelated serovar Infantis strains from other
from the reference strains (also listed in Table 1) served as positive controls for
public health institutions of northern (a susceptible strain from Para
the PCRs. Negative controls for PCRs consisted of all the reagents used for each
western Brazil (a multidrug-resistant strain from Brasilia) were used to assess the
primer pair minus the DNA template. Genomic DNA from cultures grown at
utility of PFGE as an epidemiological marker for nosocomial infections.
35°C on Mueller-Hinton agar with antimicrobials was extracted with a Puregenekit (Gentra Systems, Inc., Minneapolis, MN). The PCR mix for the detection ofresistance genes and integrons included 1.0 M of forward and reverse primers,
RESULTS AND DISCUSSION
1ϫ Taq polymerase buffer, 1.5 mM MgCl2, 200 M of each deoxynucleotide
Many researchers are successfully using PFGE to investigate
(dATP, dCTP, dGTP, and dTTP) (Gibco BRL, Burlington, Ontario), 0.025 U/lTaq polymerase (Gibco BRL, Burlington, Ontario), and approximately 1 g of
the epidemiologies of strains involved in outbreaks caused by
template DNA. Amplification conditions for all of the PCRs, except for integron
beta-lactamase- and ESBL-producing bacteria (8). The PFGE
and blaTEM amplification, were 1 cycle at 94°C for 5 min and 35 cycles for 94°C
analysis of the 35 serovar Infantis strains resulted in five PFGE
for 1 min, 55°C for 1 min, and 72°C for 1 min 30 s. An annealing temperature of
restriction fragment profiles (Fig. 1 and 2). The comparative
48°C was used for the amplification of blaTEM. Integron amplification involved 1cycle at 94°C for 12 min and 35 cycles at 94°C for 1 min, 55°C for 1 min, and 72°C
evaluation of the PFGE profiles yielded four fragment patterns
for 5 min. PCR products were analyzed by gel electrophoresis in a 1% agarose
(A1, A3, A4, and A5) for HA isolates. Three HB and five HC
TABLE 2. Antimicrobial resistance and PFGE profiles for serovar Infantis strains isolated between 1996 and 2001
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, FEP)
ACSSuTTmKG (ATM, CEF, CXM, CTX, CRO, FEP)
ACSSuTTmG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSSuTTmG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ASSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
ACSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
a Cephalosporin and aztreonam resistance profiles are shown in parentheses. A, ampicillin; C, chloramphenicol; S, streptomycin; Su, sulfamethoxazole; T,
tetracycline; Tm, trimethoprim; K, kanamycin; G, gentamicin. b PHL, Public Health Laboratory (Brası´lia, Brazil); IEC, Evandro Chagas Institute (Para, Brazil).
isolates had the PFGE profile A1 that was also encountered in
ACSSuTTmKG, etc. [Table 2]) had similar PFGE profiles and
18 HA strains. The three HD strains (PFGE profile A2), iso-
were isolated from 1996 to 2001 from patients in different
lated in 2001, showed 95% similarity to PFGE profiles A1 and
hospitals. The high prevalence of resistance to these particular
A3 (Fig. 1). Macro-restriction fragment patterns of strains
antimicrobials may be due to selective pressure since these
from northern (PFGE profile B) and midwestern (PFGE pro-
antimicrobials, with the exception of kanamycin and strepto-
file C) regions of Brazil were completely different from those
mycin, are among the agents most often prescribed in these
of strains from Rio de Janeiro, Brazil.
hospitals. Resistance to kanamycin and streptomycin, however,
The A1 profile, found in 26 strains isolated from 1996 to
may have been acquired through horizontal gene transfer since
2001 in HA, HB, and HC, was considered to be the PFGE
aminoglycoside resistance genes are often found on plasmids
profile associated with the MDR serovar Infantis outbreaks.
and transposons that encode resistance determinants for other
The PFGE patterns of serovar Infantis strains were then clas-
classes of antimicrobials (34, 36). Tetracycline resistance
sified according to their similarities to the outbreak pattern.
(97.2%) and aztreonam resistance (96.1%) were also common
Patterns that differed from the outbreak pattern by two frag-
among the multidrug-resistant strains. It is not surprising that
ments (Յ90%) were considered to be subtypes. A variation of
the four hospitals involved in this study experienced great
two to three fragments in a PFGE profile can occur when
difficulties in deciding which antimicrobials to use for treat-
strains are cultured repeatedly or isolated multiple times from
ment. The implementation of effective screening methods for
the same patient (33). Those patterns that differed by at least
the detection of beta-lactamases and ESBLs as well as the
four fragments were classified as unrelated types by consider-
establishment of surveillance programs became key factors in
ing that they derived from two genetic events and their isolat-
the control of hospital outbreaks (16).
PCR detection of resistance genes in nine isolates resistant
The susceptibility profiles of serovar Infantis are shown in
to five classes of antimicrobials, represented by ampicillin,
Table 2. All of the strains were susceptible to carbapenem
chloramphenicol, streptomycin, sulfamethoxazole, and tetracy-
(imipenem), ciprofloxacin, nalidixic acid, and cephamycin
cline, showed that all of the strains with the ACSSuTTmKG
(cefoxitin). All of the strains, except for one, were resistant to
, catI, aadA1, sulI, sulII, and
ampicillin, and most were resistant to cephalosporins (includ-
tet(D) resistance genes and an integron containing an
ing extended spectrum). It is interesting that strains resistant
aac(6Ј)-Iq gene cassette that codes for amikacin resistance
to the highest number of antimicrobials (resistance profile
(Table 3). The only variation among these strains was the
GENE PROFILES OF SALMONELLA ENTERICA SEROVAR INFANTIS
TABLE 3. Antimicrobial resistance genes detected in multidrug-resistant serovar Infantis strains
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
, catI, aadA1, sulI/II, tet(D)
ACSSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
, catI, aadA1/A2, sulI/II, tet(D)
ACSSuTTmKG (ATM, CEF, CXM, CTX, CAZ, FEP)
, catI, aadA1, sulI/II, tet(D)
ACSSuTTmKG (ATM, CEF, CXM, CTX, CRO, FEP)
, catI, aadA1/A2, sulI/II, tet(D)
ACSuTTmKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
, catI, aadA1/A2, sulI/II, tet(D)
ASSuTKG (ATM, CEF, CXM, CAZ, CTX, CRO, FEP)
, aadA1/A2, sulII, tet(D)
a Cephalosporin and aztreonam resistance profiles are shown in parentheses. A, ampicillin; C, chloramphenicol; S, streptomycin; Su, sulfamethoxazole; T,
tetracycline; Tm, trimethoprim; K, kanamycin; G, gentamicin. b The integron cassette size was 1,269 bp. c “aadA1/A2” or “sulI/II” indicates the presence of both aadA1 and aadA2 or both sulI and sulII, homologous genes, respectively, within a strain. d This strain was intermediately resistant to streptomycin.
presence or absence of the streptomycin/spectinomycin resis-
ysis provides evidence that the hospitals involved in this study
tance gene aadA2, also known as ant(3Ј)Ib, a variant of the
had a salmonellosis outbreak that was caused by serovar In-
gene aadA1 [ant(3Ј)Ia]. The serovar Infantis strain with the
fantis strains that shared the same phylogenetic lineage. It is
ACSuTTmKG resistance profile was intermediately resistant
important to emphasize that strains from HC were isolated in
to streptomycin and yet carried both aadA1 and aadA2. Re-
only 1996, while strains from HB were isolated in 1997 and
dundancy of resistance genes was also detected in 10 strains
1999. HA strains were isolated from 1996 to 1999. At the
carrying two sulfonamide resistance genes, sulI and sulII.
beginning of 2001, HD was informed about the characteristics
Thirty-two (91.4%) serovar Infantis strains were classified as
and clonal nature of multidrug-resistant serovar Infantis so
clavulanic-acid-inhibited ESBL-producing strains according to
that appropriate control measures could be developed and,
CLSI standards (21 from HA, 3 from HB, 5 from HC, and 3
subsequently, serovar Infantis was no longer detected in the
from HD). Twenty-two strains (62.8%) were resistant to both
hospital environment. The guidelines and rules that provide
CTX and CAZ (Table 2), which suggested the presence of at
for the planning of the National Program of Hospital Infection
least one ESBL. According to the beta-lactamase classification
Control were defined by administrative rule GM 2.616 as of 12
scheme of Bush et al. (1995), cefotaximases are class A ESBLs
May, 1998. This decree categorizes children hospitalized in
(group 2be) that generally have higher hydrolytic activities
high-risk nurseries as intensive-care patients requiring partic-
against cefotaxime than ceftazidime, while ceftazidimases (also
ular attention to infections due to multidrug-resistant patho-
group 2be ESBLs) generally hydrolyze ceftazidime more
gens (22). These patients are subjected to standard procedures
readily than cefotaxime (10). In addition, group 2be ESBLs
for controlling nosocomial infections, such as the cleaning and
inactivate not only extended-spectrum cephalosporins but also
disinfection of medical equipment, frequent hand washing,
monobactams such as aztreonam. Ten of the serovar Infantis
patient-to-patient contact precautions, and the monitoring of
strains characterized in this study were resistant to both CTX
patients’ stools for the presence of multidrug-resistant serovar
and CAZ, while only one was resistant to only CTX (Table 3).
Infantis. The best strategy for antimicrobial therapy and spe-
DNA sequencing of the amplicons obtained with bla
cific infection control measures for each patient was deter-
ers (which targeted the conserved region of TEM-related en-
zymes) revealed the presence of the non-ESBL bla
The results in this study indicate that efficient surveillance
order to determine whether more than one beta-lactamase was
programs and effective decontamination procedures must be
produced by these 11 multidrug-resistant serovar Infantisstrains, isoelectric focusing was performed (Table 4).
The pI profiles indicated the presence of beta-lactamases
TABLE 4. -Lactamase profiles detected in multidrug-resistant
with pI values of 5.4, 6.3, 6.9, and 9.0. The six strains with the
antibiogram ACSSuTTmKG (resistance profile, ATM, CEP,
CXM, CAZ, CTX, CRO, FEP) had at least four different
beta-lactamases (since there could be more than one beta-
lactamase present in a strain with the same pI value), while
another strain with the same resistance profile produced only
two types of beta-lactamases (pIs 9.0 and 5.4). This result is
significant since all seven strains are resistant to CEP, CXM,
CAZ, CTX, CRO, and FEP, indicating that resistance to those
cephalosporins requires the presence of only two types of beta-
lactamases with pI values of 5.4 and 9.0. In addition, those
seven strains are also resistant to the monobactam ATM,
which indicates, according to Bush et al., that a group 2be
ESBL is present within the strain (10). a Cephalosporin and aztreonam resistance profiles are shown in parentheses.
The presence of identical antimicrobial resistance genes and
A, ampicillin; C, chloramphenicol; S, streptomycin; Su, sulfamethoxazole; T,
the close relatedness of strains as determined by PFGE anal-
tetracycline; Tm, trimethoprim; K, kanamycin; G, gentamicin.
implemented for the prevention of nosocomial outbreaks of
integrons reveals several novel combinations of resistance genes. Antimi-
salmonellosis caused by multidrug-resistant serovar Infantis.
crob. Agents Chemother. 39:185–191.
19. Marshall, B., S. Morrissey, P. Flynn, and S. B. Levy. 1986. A new tetracy-
cline-resistant determinant, class E, isolated from Enterobacteriaceae. Gene
ACKNOWLEDGMENTS 50:111–117.
20. Marshall, B., C. Tachibana, and S. B. Levy. 1983. Frequency of tetracycline
We thank C. M. F. Reis and A. F. M. Santos (FIOCRUZ, Rio de
resistance determinant classes among lactose-fermenting coliforms. Antimi-
Janeiro, Brazil) for her collaboration on the PFGE technique and
crob. Agents Chemother. 24:835–840.
photo documentation and E. Soares and his working group
21. Mendez, B., C. Tachibana, and S. B. Levy. 1980. Heterogeneity of tetracy-
cline resistance determinants. Plasmid 3:99–108.
(FIOCRUZ, Rio de Janeiro, Brazil), who provided assistance and
22. Ministe´rio da Sau ´de. 1998. Portaria n° 2.616, de 12 de maio de 1998. Dispo
This work was supported by grants from the Oswaldo Cruz Institute
Pos-Graduation/FIOCRUZ-Rio de Janeiro and National Council for Sci-
23. Mulvey, M. R., D. A. Boyd, L. Baker, O. Mykytczuk, E. M. Reis, M. D. Asensi,
entific and Technological Development (CNPq), Brazil. O. Mykytczuk’s
D. P. Rodrigues, and L. K. Ng. 2004. Characterization of a Salmonella
student stipend was from the National Microbiology Laboratory and the
enterica serovar Agona strain harbouring a class 1 integron containing novel
University of Manitoba, Winnipeg, Manitoba, Canada.
OXA-type -lactamase (blaOXA-53) and 6Ј-N-aminoglycoside acetyltrans- ferase genes [aac(6Ј)-I30]. J. Antimicrob. Chemother. 54:354–359.
24. Murray, I. A., J. V. Martinez-Suarez, T. J. Close, and W. V. Shaw. 1990. REFERENCES
Nucleotide sequences of genes encoding the type II chloramphenicol acetyl-
1. Ageˆncia Nacional de Vigila ˆncia Sanita ´ria (ANVISA). 2005. Manual de pro-
transferases of Escherichia coli and Haemophilus influenzae, which are sen-
´sicos em microbiologia clı´nica para o controle de infecc¸a
sitive to inhibition by thiol-reactive reagents. Biochem. J. 272:505–510.
hospitalar. [Online.] http://www.hu.usp.br/arquivos/Manualccih_2005.pdf.
25. Murray, I. A., A. R. Hawkins, J. W. Keyte, and W. V. Shaw. 1988. Nucleotide
2. Alton, N. K., and D. Vapnek. 1979. Nucleotide sequence analysis of the
sequence analysis of overexpression of the gene encoding a type III chlor-
chloramphenicol resistance transposon Tn9. Nature 282:864–869.
amphenicol acetyltransferase. Biochem. J. 252:173–179.
3. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.
26. NCCLS. 2004. Performance standards for antimicrobial susceptibility test-
Basic local alignment search tool. J. Mol. Biol. 215:403–410.
ing; 30th informational supplement. NCCLS document no. M100–S14, vol.
4. Altschul, S. F., T. L. Madden, A. A. Scha ¨ffer, J. Zhang, Z. Zhang, W. Miller, and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation
27. Ng, L.-K., M. R. Mulvey, I. Martin, G. A. Peters, and W. Johnson. 1999.
of protein database search programs. Nucleic Acids Res. 25:3389–3402.
Genetic characterization of antimicrobial resistance in Canadian isolates of
5. Angulo, F. J., K. R. Johnson, R. V. Tauxe, and M. L. Cohen. 2000. Origins Salmonella serovar Typhimurium DT104. Antimicrob. Agents Chemother.
and consequences of antimicrobial-resistant nontyphoidal Salmonella: impli-
43:3018–3021.
cations for the use of fluoroquinolones in food animals. Microb. Drug Resist.
28. Olsen, S. J., R. Bishop, F. W. Brenner, T. H. Roels, N. Bean, R. V. Tauxe, and 6:77–83. L. Slutsker. 2001. The changing epidemiology of Salmonella: trends in se-
6. Asensi, M. D., and E. Hofer. 1994. Serovars and multiple drug resistant
rotype isolated from humans in the United States, 1987-1997. J. Infect. Dis. Salmonella sp. isolated from children in Rio de Janeiro – Brazil. Rev. Mi-
183:753–761.
˜o Paulo 25:149–153.
29. Persing, D. H., T. F. Smith, F. C. Tenover, and T. J. White (ed.). 1993.
7. Bito, A., and M. Susani. 1994. Revised analysis of aadA2 gene of plasmid
Diagnostic molecular microbiology: principles and applications. American
pSa. Antimicrob. Agents Chemother. 38:1172–1175.
Society for Microbiology, Washington, D.C.
8. Bradford, P. A. 2001. Extended-spectrum beta-lactamase in the 21st century:
29a.Poppof, M. Y. 2001. Antigenic formulas of the Salmonella serovars. WHO
characterization, epidemiology, and detection of this important resistance
Collaborating Center for Reference and Research for Salmonella Serovars,
threat. Clin. Microbiol. Rev. 14:933–951.
9. Bradley, D. E., D. E. Taylor, and D. R. Cohen. 1980. Specification of surface
30. Pessoa-Silva, C. L., C. M. Toscano, B. M. Moreira, A. L. Santos, A. C. Frota,
mating systems among conjugative drug resistance plasmids in EscherichiaC. A. Solari, E. L. Amorim, M. da Gloria S. Carvalho, L. M. Teixeira, and coli K-12. J. Bacteriol. 143:1400–1470. W. R. Jarvis. 2002. Infection due to extended-spectrum beta-lactamase-
10. Bush, K., G. A. Jacoby, and A. A. Medeiros. 1995. A functional classification
producing Salmonella enterica subsp. enterica serovar Infantis in a neonatal
scheme for -lactamases and its correlation with molecular structure. Anti-
unit. J. Pediatr. 141:381–387.
microb. Agents Chemother. 39:1211–1233.
31. Rådstro ¨m, P., and G. Swedberg. 1988. RSF1010 and a conjugative plasmid
11. Chiappini, E. L., L. Galli. P. Pecile, A. Vierucci, and M. de Martino. 2002.
contain sulII, one of two known genes for plasmid-borne sulfonamide resis-
Results of a 5-year prospective surveillance study of antibiotic resistance
tance dihydropteroate synthase. Antimicrob. Agents Chemother. 32:1684–
among Salmonella enterica isolates and ceftriaxone therapy among children
hospitalized for acute diarrhea. Clin. Ther. 24:1585–1594.
32. Sundstro ¨m, L., and O. Sko ¨ld. 1990. The dhfrI trimethoprim resistance gene
12. Costa, G. A., and E. Hofer. 1972. Isolamento e identificac¸a
can be found at specific sites in other genetic surroundings. Antimicrob.
´rias, p. 120. Instituto Oswaldo Cruz – FIOCRUZ, Rio de Janeiro, Brazil.
Agents Chemother. 34:642–650.
13. De Moraes, B. A., C. A. N. Cravo, M. M. Loureiro, C. A. Solari, and M. D.
33. Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E. Murray, Asensi. 2000. Epidemiological analysis of bacterial strains involved in hos- D. H. Persing, and B. Swaminathan. 1995. Interpreting chromosomal DNA
pital infection in a university hospital from Brazil. Rev. Inst. Med. Trop. Sa
restriction patterns produced by pulsed-field gel electrophoresis: criteria for
Paulo 42:201–207.
bacterial strain typing. J. Clin. Microbiol. 33:2233–2239.
14. Edelstain, M., and L. Stratchounski. 1998. Development of single-strand
34. Tolmasky, M. E., R. M. Chamorro, J. H. Crosa, and P. M. Marini. 1988.
conformational polymorphism (SSCP) PCR method for discriminatory de-
Transposon-mediated amikacin resistance in Klebsiella pneumoniae. Antimi-
tection of genes coding for TEM-family -lactamases. Program Abstr. 38th
crob. Agents Chemother. 32:1416–1420.
Intersci. Conf. Antimicrob. Agents Chemother., poster E-96.
35. Winokur, P. L., A. Brueggemann, D. L. DeSalvo, L. Hoffmann, M. D. Apley,
15. Hansen, L. M., L. M. McMurry, S. B. Levy, and D. C. Hirsh. 1993. A new E. K. Uhlenhopp, M. A. Pfaller, and G. V. Doern. 2000. Animal and human
tetracycline resistance determinant, Tet H, from Pasteurella multocida spec-
multidrug-resistant, cephalosporin-resistant Salmonella isolates expressing a
ifying active efflux of tetracycline. Antimicrob. Agents Chemother. 37:2699–
plasmid-mediated CMY-2 AmpC beta-lactamase. Antimicrob. Agents Che-
mother. 44:2777–2783.
16. Kaye, K. S., H. S. Fraimow, and E. Abrutyn. 2000. Pathogens resistant to
36. Vakulenko, S. B., and S. Mobashery. 2003. Versatility of aminoglycosides
antimicrobial agents—epidemiology, molecular mechanisms and clinical
and prospects for their future. Clin. Microbiol. Rev. 16:430–450.
management. Infect. Dis. Clin. N. Am. 14:293–317.
37. Zhao, J., and T. Aoki. 1992. Nucleotide sequence analysis of the class G
tetracycline resistance determinant from Vibrio anguillarum. Microbiol. Im-
18. Le´ve`sque, C., L. Piche´, C. Larose, and P. H. Roy. 1995. PCR mapping of
munol. 36:1051–1060.
Facelift Instructions This handout provides information on how to prepare for your facelift operation and what to expect as you are healing after your operation. Please read the handout carefully. Feel free to ask questions at any time. Important information about preparing for your facelift operation: 1. Do not take products containing aspirin, ibuprofen (Advil, Motrin),Vitamin E or
R. v. Khan Her Majesty the Queen, and Sayeed Khan W.B. Trafford J. Criminal law -- Offences -- Sexual offences -- Rape or sexual assault -- Consent -- The accused,charged with sexual assault, was found not guilty. Trial of the accused, who was deaf, charged with sexual assault -- Two police officers observed theaccused and the 18-year-old complainant kissing in a phone booth at a sub

 GENE PROFILES OF SALMONELLA ENTERICA SEROVAR INFANTIS
FIG. 2. PFGE macro-restriction fragment polymorphism.
GENE PROFILES OF SALMONELLA ENTERICA SEROVAR INFANTIS
FIG. 2. PFGE macro-restriction fragment polymorphism.