John A. E. Gibson á Claude Belzile á Warwick F. VincentFarthest north lake and fjord populations of calanoid copepods
Limnocalanus macrurus and Drepanopus bungei
Accepted: 21 October 2000 / Published online: 18 January2001
Abstract The zooplankton assemblages of Lake A and since that time (Jeries et al. 1984; Retelle 1986; Ludlam
Disraeli Fjord, northern Ellesmere Island (83°N, 75°W), 1996). Little attention, however, has been given to the
were surveyed in early summer 1999. In permanently ice- biological limnologyof this lake. NearbyDisraeli Fjord
covered Lake A, two glacial relict calanoid copepod has been sampled sporadicallysince 1967, and previous
species (Drepanopus bungei and Limnocalanus macrurus) collections of zooplankton have revealed the presence
were found in the top 30 m. All developmental stages of of Drepanopus bungei (Bowman and Long 1968). Given
the more abundant D. bungei were present, whereas only their extreme location at the northern limit of North
adults of L. macrurus were found. Analysis of gut con- America, the species composition at these sites is of
tents showed that L. macrurus preyed upon the smaller special interest for biogeographical analyses. In addition
species. A net tow sample of zooplankton from Disraeli to the zooplankton collections, we also undertook a
Fjord was mainlycomposed of D. bungei and L. macru- limnological analysis of these sites to provide back-
rus, along with two marine cyclopoid copepods (Oncaea ground information on habitat characteristics.
borealis and Oithona similis). These two zooplankton
communities occur within unusual environments that are
stronglyin¯uenced byperennial ice and snow. Theywill Materials and methods
be subject to major habitat disruption should the current
warming trends continue in the north polar region.
Lake A (83°00¢N, 75°30¢W; Fig. 1), and Disraeli Fjord (82°50¢N,
73°40¢W; Fig. 1) are located on the northern coast of Ellesmere
Island. Lake A is meromictic (permanentlystrati®ed) and has a
maximum depth of >115 m, a surface area of 4.9 km2, an
apparentlyperennial ice cover up to 2 m in thickness, and a
catchment area of 37 km2 containing no glaciers. It was formed
after the last ice age when isostatic uplift of northern Ellesmere
The distribution of copepods in the Arctic Ocean and Island trapped pockets of seawater in a pre-existing depression
arctic coastal waters is well documented (summarized (Lyons and Mielke 1973). Disraeli Fjord is a strati®ed, 45-km-long
in Mauchline 1998; Thibault et al. 1999); however, fjord that is presentlydammed bythe Ward Hunt Ice Shelf. At time
information concerning lake and fjord populations at of sampling, the ice cover on the fjord was 2.4 m.
Water column measurements were made in Lake A during the
extreme latitudes in the circumpolar Arctic is still lim- ®rst week of June 1999 and Disraeli Fjord was pro®led on 9 June
ited. In the present study, we examined the zooplankton 1999. Temperature, salinityand dissolved oxygen were measured
assemblages at the northern limit of these habitat types: using a Hydrolab Surveyor 3 pro®ler. Estimates of phytoplankton
perennial ice-covered Lake A and ice-dammed Disraeli biomass were made bymeasuring chlorophyll a (Chl a) concen-
tration. Water was sampled at 1-m intervals in the oxic zone in
Fjord. Both sites are located at latitude 83°N in the Lake A and at 1- to 10-m intervals in Disraeli Fjord. Sampling was
made with a 2-l Kemmerer bottle and 250-ml subsamples were
Lake A was ®rst investigated in 1969 (Hattersley- ®ltered through GF/F glass ®ber ®lters. Filters were kept frozen
Smith et al. 1970), and there have been occasional visits and pigments were extracted within less than a month, using
boiling ethanol according to Nusch (1980). Fluorescence was
measured with a Sequoia-Turner Model 450 ¯uorometer, with
correction for phaeopigments using the equations of Jereyand
P. Van Hove (&) á K. M. Swadling á J. A. E. Gibson
Zooplankton was sampled from Lake A on 8 June 1999 and
Centre d'eÂtudes nordiques, Universite Laval,
Disraeli Fjord on 9 June 1999. At Lake A, a hole was cut through
the ice and was kept open for 3 days before zooplankton sam-
pling. A 1-m-long conical plankton net (mesh size 100 lm; mouth
diameter 20 cm) was used to sample the zooplankton. Duplicate
net tows were made sequentiallyat the following depths: 2.5, 5,
Fig. 1 Location of Lake A and Disraeli Fjord on northern Ellesmere Island, Canadian high Arctic
7.5, 10, 12.5, 15, 17.5, 20, 25, and 30 m. The net was towed by
hand verticallyat approximately1 m/s from a given depth to the
surface. In Disraeli Fjord, logistical constraints allowed us onlya
single tow from the brackish-water layer, from 30 m to the sur-
face. All samples were preserved with ®nal concentrations of 0.2%
glutaraldehyde and 0.02% formaldehyde, a preservative used for
phytoplankton studies but which gave very good preservation of
zooplankton. The specimens were identi®ed and enumerated using
a binocular microscope (´32 magni®cation). For each species,
copepodite stages were counted separately, but nauplii stages were
pooled. The volume ®ltered bythe plankton net was determined
bymultiplying the mouth area bythe depth of sampling. To
calculate zooplankton densities, a ®ltration eciencyof 100% was
assumed (Tranter and Smith 1968), and the dierences between
numbers in each tow were used to calculate the stratum density.
The stomach contents of several adults of Limnocalanus macrurus
and D. bungei were determined bydissecting and mounting their
intestines on a glass microscope slide and examining them at ´400
Disraeli Fjord and Lake A were both highlystrati®ed
(Figs. 2, 3). Lake A had temperature and salinitypro®les
similar to other polar meromictic lakes (cf. Gibson
1999), with a temperature maximum below the surface Fig. 2 Water column properties of Lake A (8 June 1999)
and a gradual halocline. The oxygenated zone was lim-
ited to a 13-m supersaturated low conductivitylayer values for Chl a are similar to concentrations in other
(0.26 mS/cm, 0.13&) at the surface in which Chl a lakes in the high Arctic (Lake Char 0.46±0.78 lg l±1,
concentrations ranged from 0.2 to 0.5 lg Chl a l±1. These Lake Garrow 0.04±0.40 lg l±1; Markager et al. 1999) and
in oligotrophic lakes in the McMurdo DryValleys in
Two species of calanoid copepod, Limnocalanus ma-
crurus and Drepanopus bungei, were the onlymetazoans
Disraeli Fjord had a sharp halocline at 32 m that present in the samples collected from Lake A. Theyhave
likelyre¯ected the depth of the nearbyice shelf at the a well-documented distribution over the Siberian arctic
mouth of the fjord (Vincent et al., in press). The surface shelf, particularlyin the brackish surface waters in¯u-
waters were brackish (0.68 mS/cm, 0.67&), while salin- enced byriverine input (Zenkevitch 1963; Holmquist
ityand other conditions below 32 m were similar to the 1970). These species have also been found in some
Arctic Ocean. The water column sampled was well marine localities in the Canadian Arctic Archipelago
oxygenated with temperatures close to 0 °C. Chl a (Bowman and Long 1968; Holmquist 1970; Evans and
concentrations were verylow, reaching a maximum of Grainger 1980). Table 1 shows the abundance of each
0.3 lg l±1, comparable to the lower limit of the values stage for both species, and the densityfor every5-m
measured in the Arctic Ocean (Wheeler et al. 1996).
layer is shown in Fig. 4. The two net tows made for each
depth were not signi®cantlydierent (paired t-test,
t=0.58, P=0.57). Onlythe adult stages of L. macrurus
were present in our collections, whereas D. bungei was
represented byadults, copepodites, and nauplii. Of the
copepodid stages, CIII was the most abundant.
Previous studies have shown that L. macrurus and
D. bungei both have a 1-year life-cycle, and that the
adults overwinter (Ro and Carter 1972; Evans and
Grainger 1980). The absence of the earlier stages of
L. macrurus in the present studysuggests that the
population was composed onlyof the overwintering
adults. The developmental stages of D. bungei that were
found in Lake A, along with the record of spermato-
phores attached to some females, indicate that the
population of this species was in its reproductive phase.
The large numbers of nauplii of this species are prob-
ablyderived from reproduction bythe overwintering
The enumeration data show that a high proportion of
the Lake A zooplankton population resided in the 5- to
10-m stratum, close to the oxic-anoxic interface, with a
second densitymaximum in the 15- to 20-m stratum, well
into the anoxic but non-sul®dic zone. This lower peak
mayindicate a deep population of food particles and an
area of refuge from predation. The presence of ®sh has
Fig. 3 Water column properties of Disraeli Fjord (9 June 1999)
not been determined, but observations from other lakes
Table 1 Number of individuals of each species and life-cycle stage of zooplankton found in net tows in Lake A, and size for these stages.
For the number of individuals per tow, each value is the mean of two net hauls (range in parentheses)
Table 2 Number of individuals of each species and life-cycle stages
of zooplankton in Disraeli Fjord from a 20-m tow (0.63 m3 ®ltered
absence from all tows suggests that theydid not occur
in Lake A or in the surface waters of Disraeli Fjord.
Oncaea borealis and Oithona similis, found in Disraeli
Fjord, are common constituents of the Arctic Ocean
Fig. 4 Copepod densities in Lake A, northern Ellesmere Island.
zooplankton, re¯ecting the direct contact of Disraeli
The values for each stratum were calculated from dierences
Fjord with the open sea. However, the presence of
between concentrations in tows from adjacent depths
L. macrurus and D. bungei in Disraeli Fjord sets it apart
from the Arctic Ocean, and provides an intermediate
in the area (Lake C3, R.S. Bradley, personal communi- situation between Lake A and marine assemblages.
cation; Lake Garrow, Dickman 1995) suggest that there
The environmental conditions of Lake A and Disraeli
maybe a population in the lake. It is possible that the Fjord are highlydependent on the presence of ice, which
observed distribution of copepods was in¯uenced bythe limits the amount of light, wind-induced mixing and, in
sampling hole being open for 3 days before the sampling, the case of Disraeli Fjord, acts as a dam that maintains
in particular given the low irradiances that theynormally the presence of a low-salinitysurface layer. Climate
experience beneath the snow and ice (<1% of surface change can greatlyin¯uence ice conditions and current
incident irradiance, unpublished results).
monitoring data show that the Arctic is undergoing
The gut contents of six male and four female considerable warming and ice melt at present (Hart-
L. macrurus were examined from the Lake A samples. mann et al. 2000; Vincent et al., in press). If these trends
Fragments of crustacean legs were found in several of continue, then the zooplankton assemblages recorded
the guts, and one entire nauplius of D. bungei was here will be subject to major disruption and loss of their
present in the gut of an adult male. This is evidence of unusual habitats.
predadorybehavior byL. macrurus, consistent with
previous studies (Warren 1985) and suggests that Acknowledgements We thank the Natural Sciences and Engineer-
L. macrurus could be a major controlling factor on the ing Research Council (NSERC), Fonds pour la Formation des
Chercheurs et l'Aide aÁ la Recherche (FCAR) and Centre d'eÂtudes
Drepanopus population. The gut contents of ®ve female nordiques (Universite Laval) for ®nancial support. We also thank
adult D. bungei were also examined but no recognizable the Polar Continental Shelf Project for logistical support. This is
The plankton samples collected from Disraeli Fjord
were dominated bycopepodites and adults of D. bungei
(78%) (Table 2). Adult male and female L. macrurus References
were also present, but in verylow numbers (1%). Other
organisms present in the plankton tow included the early Bowman TE, Long A (1968) Relict population of Drepanopus
copepodite stages of two species of cyclopoid copepods,
bungei and Limnocalanus macrurus grimaldii (Copepoda: Ca-
Oithona similis (3%) and Oncaea borealis (4%), and
lanoida) from Ellesmere Island, N.W.T. Arctic 21: 172±180
Cairns AA (1967) The zooplankton of TanquaryFjord, Ellesmere
Island, with special reference to calanoid copepods. J Fish Res
The zooplankton species of Lake A are not found in
the coastal waters of northern Ellesmere, or elsewhere in Dickman M (1995) An isolated population of fourhorn sculpins
the central Arctic Ocean (Grainger 1964). The marine
(Myxocephalus quadricornis, familyCottidae) in a hypersaline
copepod assemblage in nearbyNansen Sound (between
high Arctic Canadian lake. Hydrobiologia 312: 27±35
Evans MS, Grainger EH (1980) Zooplankton in a Canadian Arctic
Ellesmere and Axel Heiberg Islands) is dominated by
estuary. In: Kennedy VS (ed) Estuarine research. Academic
Calanus spp. (Cairns 1967), which in general dominate
zooplankton assemblages of the Arctic Ocean (Grainger Gibson JAE (1999) The meromictic lakes and strati®ed marine
1964; Thibault et al. 1999) Although large-bodied zoo-
basins of the Vestfold Hills, East Antarctica. Antarct Sci 11:
plankton such as Calanus can be under-represented in
Grainger EH (1964) Zooplankton from the Arctic Ocean and
net hauls of the type obtained here, their complete
adjacent Canadian waters. J Fish Res Board Can 22: 543±564
Hartmann DL, Wallace JM, Limpasuvan V, Thompson DWJ,
Nusch EA (1980) Comparison of dierent methods for chlorophyll
Holton JR (2000) Can ozone depletion and global warming
and phaeopigment determination. Arch Hydrobiol Beih Ergebn
interact to produce rapid climate change? Proc Natl Acad Sci
Retelle MJ (1986) Stratigraphyand sedimentologyof coastal
Hattersley-Smith G, Keys JE, Serson H, Mielke JE (1970) Density
lacustrine basins, Northeastern Ellesmere Island, N.W.T.
strati®ed lakes in Northern Ellesmere Island. Nature 225:
Ro JC, Carter JHC (1972) Life cycle and seasonal abundance of
Holmquist C (1970) The genus Limnocalanus (Crustacea, Cope-
the copepod Limnocalanus macrurus Sars in a high arctic lake.
poda). Z Zool Syst Evolutionsforsch 8: 273±296
Thibault D, Head EJH, Wheeler PA (1999) Mesozooplankton in
¯uorimetric equations in common use in oceanography. In:
the Arctic Ocean in summer. Deep Sea Res I 46: 1391±1415
JereySW, Mantoura RFC, Wright SW (eds) Phytoplankton
Tranter DJ, Smith PE (1968) Filtration performance. In: Tranter
pigments in oceanography: guidelines to modern methods.
DJ, Fraser JH (eds) Zooplankton sampling (monographs on
oceanographic methodology2). UNESCO, Paris, pp 27±56
Jeries MO, Krouse HR, Shakur MA, Harris SA (1984) Isotope
Vincent WF (1987) Antarctic limnology. In: Viner AB (ed) Inland
geochemistryof strati®ed Lake ``A'', Ellesmere Island, NWT,
waters of New Zealand. DSIR Science Information Publishing
Ludlam SD (1996) The comparative limnologyof high arctic,
Vincent WF, Gibson JAE, Jeries MO (in press) Ice shelf collapse,
coastal, meromictic lakes. J Paleolimnol 16: 111±131
climate change and habitat loss in the Canadian High Arctic.
Lyons JB, Mielke JE (1973) Holocene history of a portion of
northernmost Ellesmere Island. Arctic 26: 314±323
Warren GJ (1985) Predaceous feeding habits of Limnocalanus
Markager S, Vincent WF, Tang EPY (1999) Carbon ®xation
toplankton in high Arctic lakes: implications of
Wheeler PA, Gosselin M, Sherr E, Thibault D, Kirchman DL,
low temperature for photosynthesis. Limnol Oceanogr 44:
Benner R, Whitledge TE (1996) Active cycling of organic
carbon in the central Arctic Ocean. Nature 380: 697±699
Mauchline J (1998) Advances in marine biology, vol 33. The
Zenkevitch L (1963) Biologyof the seas of the U.S.S.R.
biologyof calanoid copepods. Academic Press, San Diego
Extrait du bulletin de liaison "Le Défi n°16, décembre 2003, LHFB. MALADIE DE HUNTINGTON : RECHERCHE FONDAMENTALE ET MODELES ANIMAUX Raphael Hourez, Laboratoire de Neurophysiologie, ULB-Erasme, [email protected] et David Blum, Laboratoire de Neurochirurgie Expérimentale/IRIBHM, ULB-Erasme, [email protected] Partie 1 : Mécanismes de toxicité de la Huntingtine mutée Du g
Examples of Qualifying Medical Expenses Medical Flexible Spending Account (Defined in Section 213 of the Internal Revenue Code) Definition Prescribed and OTC drugs qualify however, the cost of regular skin care (face creams etc.) do not qualify (e.g. Retin-A can be used to treat acne and wrinkles), a physician’s letter of medical necessity would be required. Medical expenses
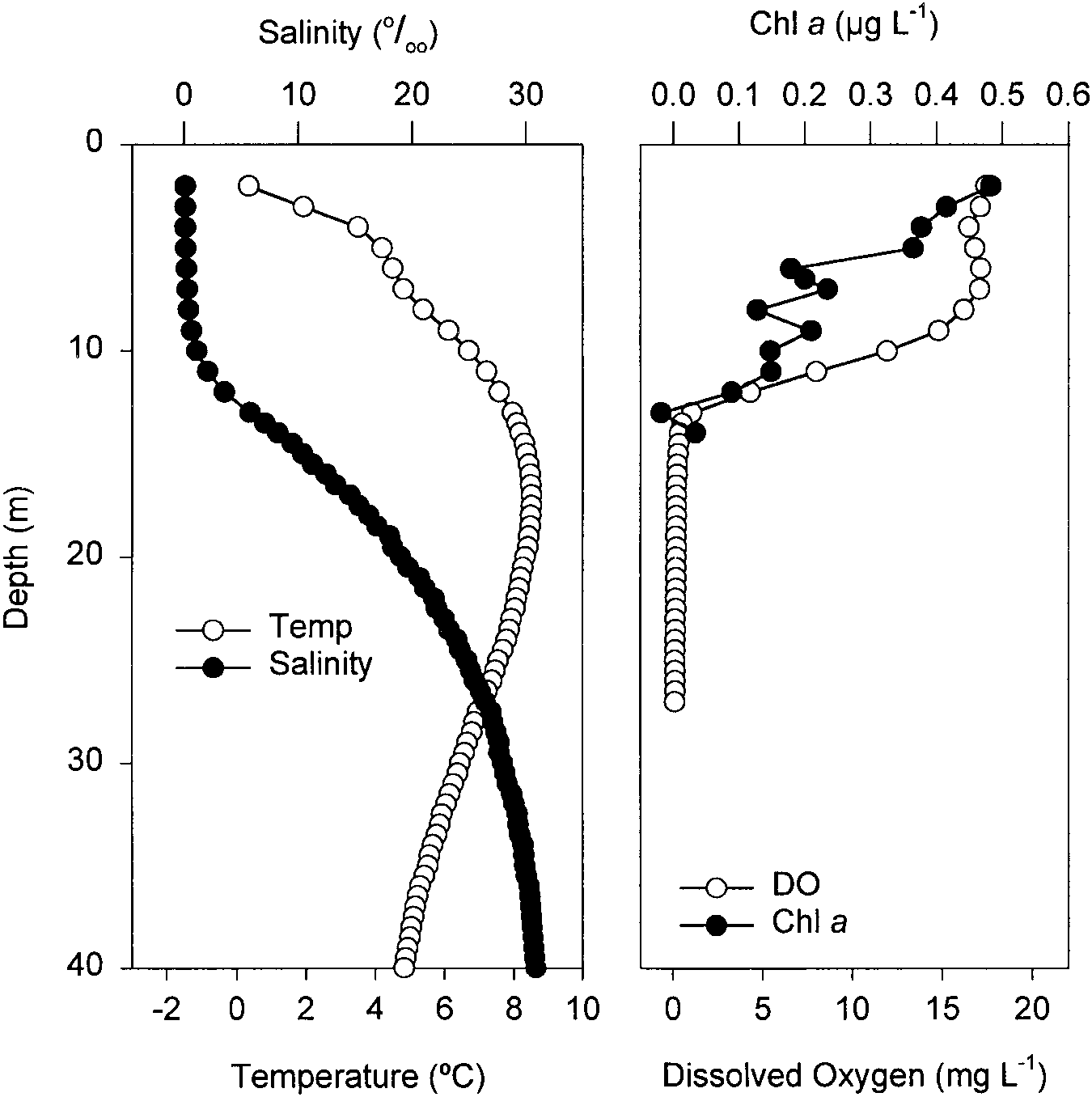
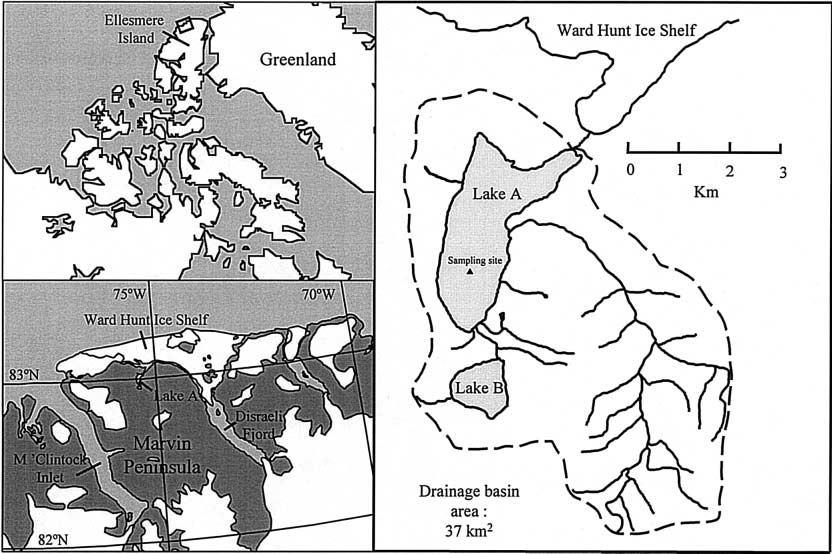 Fig. 1 Location of Lake A and Disraeli Fjord on northern Ellesmere Island, Canadian high Arctic
7.5, 10, 12.5, 15, 17.5, 20, 25, and 30 m. The net was towed by
hand verticallyat approximately1 m/s from a given depth to the
surface. In Disraeli Fjord, logistical constraints allowed us onlya
single tow from the brackish-water layer, from 30 m to the sur-
face. All samples were preserved with ®nal concentrations of 0.2%
glutaraldehyde and 0.02% formaldehyde, a preservative used for
phytoplankton studies but which gave very good preservation of
zooplankton. The specimens were identi®ed and enumerated using
a binocular microscope (´32 magni®cation). For each species,
copepodite stages were counted separately, but nauplii stages were
pooled. The volume ®ltered bythe plankton net was determined
bymultiplying the mouth area bythe depth of sampling. To
calculate zooplankton densities, a ®ltration eciencyof 100% was
assumed (Tranter and Smith 1968), and the dierences between
numbers in each tow were used to calculate the stratum density.
Fig. 1 Location of Lake A and Disraeli Fjord on northern Ellesmere Island, Canadian high Arctic
7.5, 10, 12.5, 15, 17.5, 20, 25, and 30 m. The net was towed by
hand verticallyat approximately1 m/s from a given depth to the
surface. In Disraeli Fjord, logistical constraints allowed us onlya
single tow from the brackish-water layer, from 30 m to the sur-
face. All samples were preserved with ®nal concentrations of 0.2%
glutaraldehyde and 0.02% formaldehyde, a preservative used for
phytoplankton studies but which gave very good preservation of
zooplankton. The specimens were identi®ed and enumerated using
a binocular microscope (´32 magni®cation). For each species,
copepodite stages were counted separately, but nauplii stages were
pooled. The volume ®ltered bythe plankton net was determined
bymultiplying the mouth area bythe depth of sampling. To
calculate zooplankton densities, a ®ltration eciencyof 100% was
assumed (Tranter and Smith 1968), and the dierences between
numbers in each tow were used to calculate the stratum density.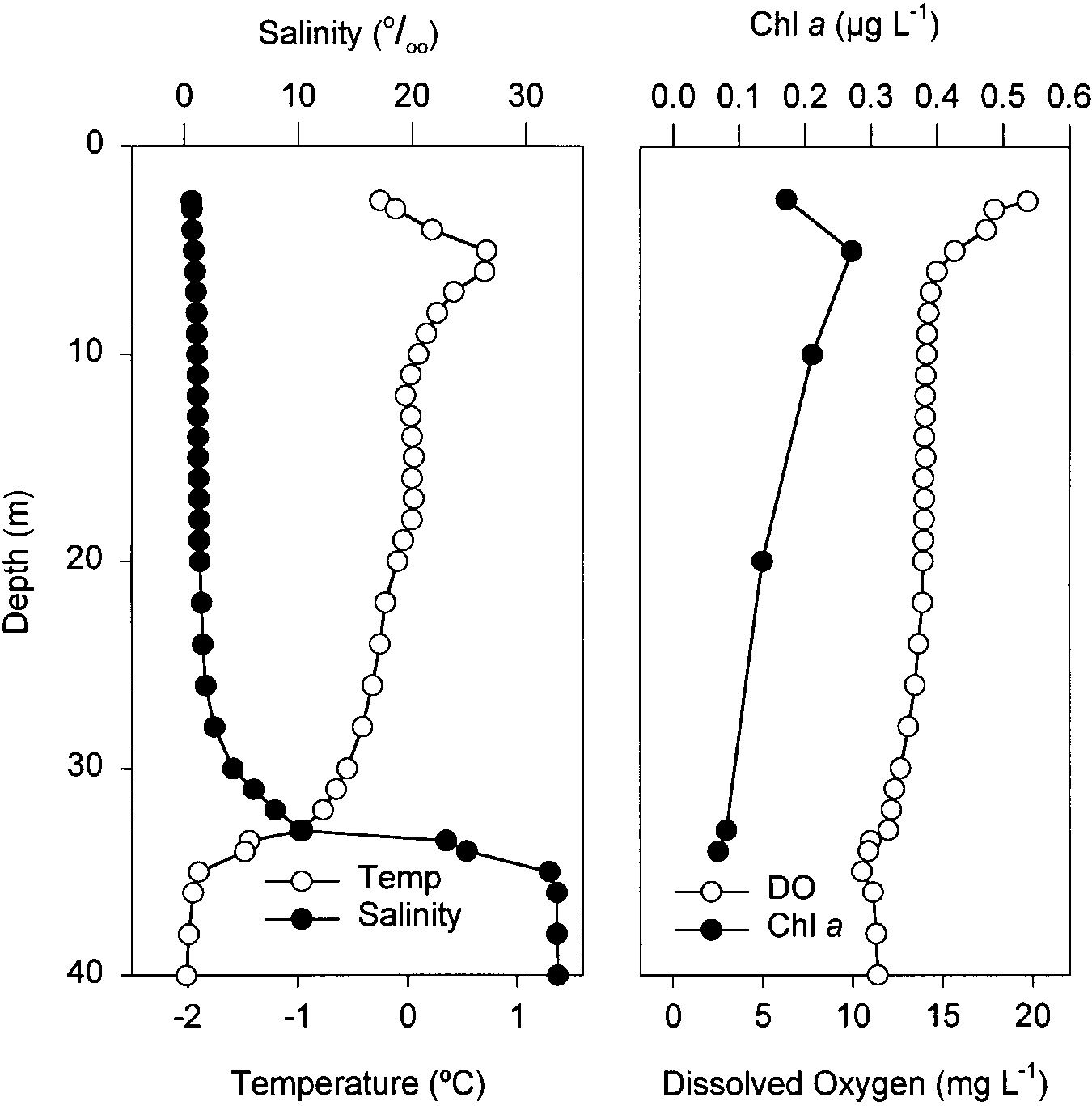 in oligotrophic lakes in the McMurdo DryValleys in
Two species of calanoid copepod, Limnocalanus ma-
crurus and Drepanopus bungei, were the onlymetazoans
Disraeli Fjord had a sharp halocline at 32 m that present in the samples collected from Lake A. Theyhave
likelyre¯ected the depth of the nearbyice shelf at the a well-documented distribution over the Siberian arctic
mouth of the fjord (Vincent et al., in press). The surface shelf, particularlyin the brackish surface waters in¯u-
waters were brackish (0.68 mS/cm, 0.67&), while salin- enced byriverine input (Zenkevitch 1963; Holmquist
ityand other conditions below 32 m were similar to the 1970). These species have also been found in some
Arctic Ocean. The water column sampled was well marine localities in the Canadian Arctic Archipelago
oxygenated with temperatures close to 0 °C. Chl a (Bowman and Long 1968; Holmquist 1970; Evans and
concentrations were verylow, reaching a maximum of Grainger 1980). Table 1 shows the abundance of each
0.3 lg l±1, comparable to the lower limit of the values stage for both species, and the densityfor every5-m
measured in the Arctic Ocean (Wheeler et al. 1996).
in oligotrophic lakes in the McMurdo DryValleys in
Two species of calanoid copepod, Limnocalanus ma-
crurus and Drepanopus bungei, were the onlymetazoans
Disraeli Fjord had a sharp halocline at 32 m that present in the samples collected from Lake A. Theyhave
likelyre¯ected the depth of the nearbyice shelf at the a well-documented distribution over the Siberian arctic
mouth of the fjord (Vincent et al., in press). The surface shelf, particularlyin the brackish surface waters in¯u-
waters were brackish (0.68 mS/cm, 0.67&), while salin- enced byriverine input (Zenkevitch 1963; Holmquist
ityand other conditions below 32 m were similar to the 1970). These species have also been found in some
Arctic Ocean. The water column sampled was well marine localities in the Canadian Arctic Archipelago
oxygenated with temperatures close to 0 °C. Chl a (Bowman and Long 1968; Holmquist 1970; Evans and
concentrations were verylow, reaching a maximum of Grainger 1980). Table 1 shows the abundance of each
0.3 lg l±1, comparable to the lower limit of the values stage for both species, and the densityfor every5-m
measured in the Arctic Ocean (Wheeler et al. 1996).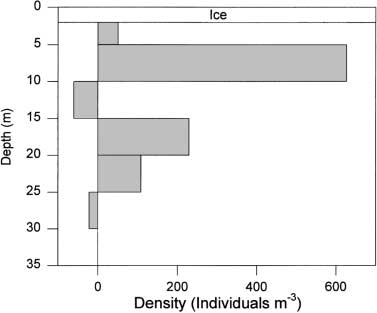 Table 2 Number of individuals of each species and life-cycle stages
of zooplankton in Disraeli Fjord from a 20-m tow (0.63 m3 ®ltered
absence from all tows suggests that theydid not occur
in Lake A or in the surface waters of Disraeli Fjord.
Table 2 Number of individuals of each species and life-cycle stages
of zooplankton in Disraeli Fjord from a 20-m tow (0.63 m3 ®ltered
absence from all tows suggests that theydid not occur
in Lake A or in the surface waters of Disraeli Fjord.