Die Struktur von Tadalafil erlaubt eine selektive Bindung an die Bindungsstelle der PDE5 und minimiert gleichzeitig die Interaktion mit PDE6, was visuelle Nebenwirkungen einschränkt. Seine Verteilung im Organismus erfolgt breit, wobei das Verteilungsvolumen etwa 63 Liter beträgt. Über 90 % des Wirkstoffs sind an Plasmaproteine gebunden. Die Wirkung bleibt unabhängig von der Nahrungsaufnahme konstant. Der Abbauweg über CYP3A4 kann durch Hemmer wie Ritonavir oder Ketoconazol verlangsamt werden, was die Plasmakonzentrationen deutlich erhöht. In diesem Kontext wird cialis 20mg preis häufig in Bezug auf pharmakokinetische Wechselwirkungen erwähnt.
Bez tytu³u-
Zastosowanie technologii interferencji RNA w medycynie
Application of RNA interference in medicine
Marta Gabryelska1, Eliza Wyszko1, Stanis³aw Nowak2, Ryszard ¯ukiel2, Jan Barciszewski1
2 z Katedry i Kliniki Neurochirurgii i Neurotraumatologii
genny RNA, np. wirusowy (18). Dlatego w³anie RNAi
Interferencja RNA nale¿y do technik anty-mRNA ada-
mo¿e funkcjonowaæ jako specyficzny stra¿nik genomu
ptowanych do celów terapeutycznych. Ma potencjalne
oraz regulator ekspresji genów (18). Zjawisko interferen-
zastosowanie w leczeniu chorób wirusowych, nowotwo-
cji RNA badano u A. thaliana, S. pombe, S. cerevisiae,
rowych, neurodegeneracyjnych i innych. Wykorzystanie
D. melanogaster, C. elegans, N. crassa, M. musculus,
RNAi zale¿y od trwa³oci wyciszenia docelowego genu,
H. sapiens oraz wielu innych organizmach (1). Mimo
³atwoci dostarczenia siRNA do chorej tkanki z zachowa-
wielu podobieñstw wystêpuj¹ drobne ró¿nice miedzy or-
niem jej aktywnoci oraz brakiem efektów ubocznych.
ganizmami. Dotycz¹ one roli tego procesu w komórkach,
Obecnie prowadzone s¹ liczne badania kliniczne spraw-
rozprzestrzeniania siê sygna³u wyciszania, obecnoci
dzaj¹ce skutecznoæ i bezpieczeñstwo terapii opartej na
okrelonych komponentów maszynerii RNAi oraz mo¿-
tej technologii. Wiele firm farmaceutycznych i biotechno-
liwoci ich wykorzystania (18, 19).
logicznych pracuje nad komercjalizacj¹ zastosowañ RNAi.
W 2001 roku przeprowadzono pierwsze eksperymenty
z wykorzystaniem siRNA w komórkach ssaków, a tak¿e
pojawi³y siê doniesienia o efektach ubocznych tej tech-
RNA interference is one of anty-mRNA techniques,
niki (62). Nie zmieni³o to jednak faktu, ¿e interferencja
adapted for a therapeutic purpose. It can be used for treat-
RNA jest bardzo obiecuj¹c¹ metod¹ wyciszania genów
ment of cancer, viral, neurodegenerative or other diseases.
ciesz¹c¹ siê wielkim zainteresowaniem zarówno w ba-
Therapeutic applications of RNAi depend on persistent gene
daniach podstawowych jak i w biotechnologicznych za-
silencing and lack of side effects. Clinical trials are in
progress, checking an effectiveness and safety of RNAi.
Many pharmaceutical and biotechnological companies are
seeking for commercial application of that technology.
Konwencjonalne metody terapeutyczne maj¹ na celu
zahamowanie syntezy lub aktywnoci biologicznej da-
S³owa kluczowe: interferencja RNA, inhibicja ekspre-
nej cz¹steczki, bia³ka strukturalnego, enzymu lub recep-
sji genów, badania kliniczne, gen docelowy, efekty uboczne
tora. Wp³yw dowolnego terapeutyku na komórkê mo¿e
Key words: RNA interference, gene epression inhibi-
dotyczyæ transkrypcji docelowego genu, etapu potran-
skrypcyjnego lub potranslacyjnego. Prze³omem w poszu-
kiwaniu nowych preparatów medycznych okaza³y siê
wykonane 30 lat temu badania antysensownych kwasów
Organizmy eukariotyczne maj¹ naturalny mechanizm
nukleinowych. Nieco póniej pojawi³y siê trójniciowe
obronny w postaci interferencji RNA (ang. RNA interfe-
kompleksy (3), rybozymy oraz aptamery i 10 lat temu in-
rence, RNAi). Jest to proces, w którym d³ugie odcinki
terferencja RNA. Systematyczne wyciszanie kolejnych
dwuniciowych RNA ulegaj¹ hydrolizie do krótkich du-
produktów genów przy pomocy RNAi mo¿e byæ pomoc-
pleksów RNA o d³ugoci 21 do 28 nukleotydów oraz in-
ne w znalezieniu genu krytycznego dla wzrostu komórek
dukuj¹ swoist¹ degradacjê jednoniciowych RNA (ang.
rakowych, a mniej istotnego dla proliferacji komórek zdro-
single stranded RNA, ssRNA), takich jak mRNA lub egzo-
wych (39). Jego identyfikacja u³atwi zaprojektowanie
Tabela 1. Charakterystyka kwasów nukleinowych wykorzystywanych do inhibicji mRNA (5). PNA ang. peptide
nucleic acid, LNA ang. locked nucleic acid.
Jednoniciowe; 16-30 nukleotydów Dwuniciowe; 19-21 nukleotydów
RNAi, które mo¿e byæ wykorzystane przeciwko dowol-
wiadczalnej ma³py lepotê, prawdopodobnie z powodu
nemu czynnikowi patologicznemu, zarówno endogen-
zbyt wysokich dawek, natomiast inny lek z tej grupy nie
nemu jak i egzogennemu. W odniesieniu do chorób o zde-
hamowa³ rozwoju nowotworu u chorych z zaawanso-
finiowanej etiologii przeprowadzono badania z wykorzy-
wanym rakiem piersi (66). Dotychczas ¿aden lek oparty
staniem RNAi in vitro i in vivo w organizmach modelo-
na katalitycznych RNA nie zosta³ wprowadzony do prak-
tyki medycznej i zatwierdzony przez FDA (ang. Food and
Interferencyjny RNA obok rybozymów, deoxyrybozy-
Drug Administration) (56). Etap badañ klinicznych osi¹-
mów i antysensownych oligonukleotydów zalicza siê do
gnê³y dotychczas preparaty Angiozyme (Faza II) oraz
technologii anty-mRNA (tab. 1). Antysensowne oligonu-
kleotydy (ang. antisense-oligodeoxynucleotides, AS-ON),
Pierwsze zastosowania RNAi wzbudzi³y wielki entu-
s¹ to ³añcuchy (10-20 nukleotydowe) komplementarne
zjazm, jakiego naukowcy nie dowiadczyli wczeniej
do docelowego mRNA (38). Wiêkszoæ z nich aktywuje
w przypadku innych metod (39). Jednak¿e jej powodze-
RNazê H, która hydrolizuje niæ RNA w utworzonych he-
nie jako nowej techniki terapeutycznej bêdzie zale¿a³o
terodupleksach DNA-RNA. Inne AS-ON, nie aktywuj¹ce
g³ównie od mo¿liwoci wprowadzenia nie zdegradowa-
RNazy H, powoduj¹ zahamowanie translacji poprzez
nego leku do chorych komórek, który musi byæ wystar-
blokowanie rozpoznania przez rybosom (38). Dotychczas
czaj¹co trwa³y, aby wyciszyæ mRNA (66). Ka¿dy lek oparty
tylko jeden lek oparty na AS-ON (Vitravene) wykorzysty-
o RNAi musi byæ zatem stabilny, selektywny, nietoksycz-
wany jest w leczeniu choroby oczu wywo³anej przez
ny, móc pokonaæ barierê w postaci b³on komórkowych
cytomegalowirusy (CMV) u pacjentów z AIDS (38, 56).
i nie powodowaæ negatywnego wp³ywu na organizm (64).
Jednak nowsza metoda leczenia AIDS HAART (ang. Hi-
ghly Active Anti-Retroviral Therapy) skutecznie zapobie-
ga zapaleniom siatkówki i popyt na lek DNA jest ograni-
Oczywistym celem ograniczenia ekspresji genów z wy-
czony (66). Innym terapeutykiem typu AS-ON jest prepa-
korzystaniem technologii RNAi s¹ wirusy stanowi¹ce po-
rat Genasense, obni¿aj¹cy poziom bia³ka Bcl-2 (56).
wa¿ne zagro¿enie dla cz³owieka, miedzy innymi wirus HIV
Rybozymy s¹ katalitycznymi cz¹steczkami RNA, któ-
(ang. human immunodeficiency virus), wirusy grypy (ang.
re swoicie hydrolizuj¹ mRNA i uniemo¿liwiaj¹ syntezê
influenza virus), wirusowego zapalenia w¹troby typu B
kodowanego przez niego bia³ka. Praktyczne zastosowa-
(ang. hepatitis B virus, HBV) i C (HCV), wirusy polio (ang.
nie rybozymów zosta³o poddane w w¹tpliwoæ, ponie-
poliovirus), brodawczaka (ang. human papilloma virus,
wa¿ rybozymowy lek na ¿ó³taczkê typu C wywo³a³ u do-
HPV), wirusy herpes (ang. herpes simplex virus, HSV),
Tabela 2. Wybrane przyk³ady inhibicji infekcji wirusowych za pomoc¹ interferencji RNA. NP bia³ko nukleokapsydu,
PA sk³adnik transkryptazy RNA wirusa grypy, bia³ko 94 kDa, U38 - polimeraza DNA, UL27 - glikoproteina, UL29
bia³ko wi¹¿¹ce DNA, UL54 polimeraza DNA, Env poliproteina otoczki wirusa, Tat transaktywator transkrypcji, Rev
bia³ko regulatorowe, Zta czynnik transkrypcyjny, TRS sekwencja reguluj¹ca transkrypcjê, Spike bia³ko otoczki,
PBMCs - ang. peripheral blood mononuclear cells, jednoj¹drzaste komórki krwi obwodowej (limfocyty, monocyty).
rotawirusy, RSV (ang. respiratory syncytial virus), cytome-
noæ takie jak specyficzne receptory komórkowe czy
galowirusy (ang. human cytomegalovirus, HCMV), wirus
czynniki transkrypcyjne (64). G³ównym problemem w le-
West Nile i wiele innych (64). Wiele badañ in vitro oraz in
czeniu chorób infekcyjnych jest opornoæ na stosowane
vivo, maj¹cych na celu inhibicjê ekspresji wirusowych
leki. Dotyczy to zw³aszcza szybko mutuj¹cych wirusów.
genów, przynios³o jednoznaczne wyniki wskazuj¹ce na
Wydaje siê zatem, ¿e inhibicja wirusa wieloma interferu-
mo¿liwoæ hamowania replikacji wirusów (tab. 2).
j¹cymi RNA jednoczenie skierowanymi przeciwko ró¿-
Jako geny docelowe u wirusów wymienia siê te zaan-
nym regionom genomu zmniejsza jego szanse na unik-
ga¿owane w replikacjê i sk³adanie cz¹stek wirusowych,
niêcie efektu RNAi poprzez spontaniczne mutacje (64).
specyficzne sekwencje ich genomów oraz transkrypty
Ludzkie wirusy wykszta³ci³y pewn¹ formê ochrony
komórkowe gospodarza u³atwiaj¹ce inwazjê lub patogen-
genomu przed efektem RNAi, opart¹ o oddzia³ywania
Tabela 3. Wybrane przyk³ady terapii chorób nowotworowych za pomoc¹ RNAi. E6, E7 regulatory replikacji wirusa,
EGFR receptor czynnika wzrostu naskórka, XIAP zwi¹zany z chromosomem X inhibitor apoptozy, K-ras regula-
tor cyklu komórkowego, COX-2 - cyclooksygenaza-2, Bcr-abl bia³aczkowy gen fuzyjny.
pomiêdzy kapsydem a genomem (58). Nukleokapsyd
apoptozy (APOP) (50). Wród wyciszanych genów znaj-
wirusa miêsaka Rousa (ang. Rous sarcoma virus) os³ania
duj¹ siê geny wirusowe, onkogeny, geny fuzyjne, geny
wprowadzany wirusowy RNA przed degradacj¹, a koñ-
opornoci wielolekowej (ang. multidrug resistance, MDR)
cowy efekt RNAi zale¿y od wybranego docelowego frag-
(32), regulatory proliferacjii i inne (tab. 3).
mentu RNA (64). Podobny rodzaj supresji RNAi jest ob-
Obecnoæ mechanizmu RNAi potwierdzono tak¿e
serwowany w przypadku wirusa RSV (ang. respiratory
u Trypanosoma brucei, Plasmodium falciparum, Toxopla-
syncytial virus), gdzie genom mo¿e byæ niedostêpny dla
sma gondii, Paramecium, Leichmania donovanii (1). Jest
siRNA z powodu utworzenia kompleksu z bia³kiem (45).
to istotne poniewa¿ pierwotniaki s¹ ród³em wielu chorób
Podobna sytuacja dotyczy innych wirusów, np. HIV. Z te-
(64). Wyzwanie dla interferencji RNA stanowi¹ tak¿e cho-
go powodu proces degradacji docelowego RNA musi roz-
roby wywo³ane przez takie czynniki jak cukrzyca, czy oty-
pocz¹æ siê w odpowiednim momencie cyklu rozwojo-
³oæ (tab. 4), choroby wywo³ane przez mutacje punktowe,
wego wirusa, kiedy jego genom bêdzie dostêpny.
choroby neurodegeneracyjne oraz zwi¹zane ze zmienn¹
Do g³ównych markerów biochemicznych zaanga¿o-
liczb¹ powtórzeñ mikrosatelitarnych. Du¿a dok³adnoæ
wanych w powstawanie nowotworów zalicza siê recep-
i skutecznoæ dzia³ania siRNA umo¿liwia leczenie chorób
tor bia³kowej kinazy tyrozynowej (ang. protein tyrosine
wywo³anych przez mutacje punktowe, insercje czy dele-
kinase, PTK), gruczolakowatej polipowatoci coli (ang.
cje zidentyfikowanych genów (18). Olbrzymim wyzwaniem
adenomatous polyposis coli, APC), onkogen Gli (ang. glio-
s¹ choroby, u których pod³o¿a le¿y alternatywny splicing,
ma-associated oncogene, Gli), fosfoinozytolow¹ kinazê
który w oko³o 70%-90% prowadzi do powstania bia³ek
3 (ang. phosphoinisitide 3-kinase, PIK3), szlak transduk-
o zmienionych funkcjach (20). Do chorób tych zaliczane
cji bia³ek SMAD, czynnik transkrypcyjny indukuj¹cy nie-
s¹ miêdzy innymi demencja czo³owo-skroniowa (ang. fron-
dotlenienie (ang. hypoxia-inducible transcription factor,
totemporal dementia), parkinsonizm zwi¹zany z chromo-
HIF), retinoblastoma (ang. retinoblastoma, Rb) oraz szlak
somem 17, niedobór hormonu wzrostu i inne (20).
Tabela 4. Terapia innych chorób za pomoc¹ RNAi; berghepain-1 i 2 proteaza cysteinowa, DHODH dehydrogenaza
dihydroorotowianu, PEPCK karboksykinaza fosfoenolo-pirogronianowa, AGRP - agouti-related peptide, SOD1 dys-
mutaza ponadtlenkowa, Htt - IT15, huntingtyna.
cz¹steczek jest ograniczone ze wzglêdu na nietrwa³y efekt
Jak podkrelono wczeniej terapeutyczne zastosowa-
wyciszenia, który zale¿y od tempa podzia³ów komórko-
nie RNAi zale¿y od trwa³oci wyciszenia, ³atwoci roz-
wych, komórki ssaków nie maj¹ systemu powielania RNAi
przestrzeniania siê siRNA w obrêbie chorej tkanki, ich ak-
(jak w przypadku C. elegans i rolin) (26).
tywnoci biologicznej, a tak¿e brak efektów ubocznych.
W badaniach procesu interferencji RNA istotna jest
siRNA uwa¿a siê za skuteczny, je¿eli w stê¿eniu 1-20 nM
odpowiednia linia komórkowa, je¿eli dowiadczenia na
prowadzi do efektywnego i specyficznego wyciszenia 90%
¿ywym organizmie s¹ utrudnione lub niemo¿liwe. Ko-
(47). Ró¿nice w efektywnoci siRNA, skutecznoci trans-
mórki takie musz¹ byæ ³atwe w hodowli, ³atwo ulegaæ
fekcji, rodzaju wykorzystanej linii komórkowej i stabilno-
transfekcji oraz posiadaæ nisk¹ wra¿liwoæ na ewentual-
ci bia³ek mog¹ wp³ywaæ na osi¹gniêty efekt RNAi (45).
ne modyfikacje chemiczne dostarczanych cz¹steczek siR-
Nale¿y pamiêtaæ, ¿e rezultaty uzyskane w badaniach in
NA. Bardzo czêsto wykorzystuje siê komórki nowotwo-
vitro nie ³atwo przek³adaj¹ siê na wyniki otrzymane in vivo.
rowe ze wzglêdu na ich ³atwe namna¿anie (tab. 5), cho-
Rolê induktora wyciszania (ang. trigger) w przypadku
cia¿ nie wszystkie typy schorzeñ s¹ reprezentowane przez
ssaków skutecznie spe³niaj¹ dwuniciowe shRNA (ang.
short hairpin RNA) lub siRNA (ang. short interfering RNA)
Przylegaj¹ce do pod³o¿a linie komórkowe, takie jak
o d³ugoci 20 do 23 nukleotydów, zawieraj¹ce niesparo-
HeLa i U2OS umo¿liwiaj¹ ³atwy, efektywny sposób wpro-
wane 2 nukleotydy przy koñcu 3' (60). U ssaków d³ugie
wadzenia cz¹steczek interferencyjnego RNA oraz szyb-
cz¹steczki dsRNA indukowa³y interferon (60). W przy-
ki, silny wzrost w dobrze zorganizowanych pojedynczych
padku innych grup organizmów oraz niektórych linii ko-
warstwach, co u³atwia obserwacjê mikroskopow¹ (15).
mórkowych (np. ES, P19) mo¿liwe jest u¿ycie dsRNA bez
Wtórne linie komórkowe nie s¹ odpowiednikami ludz-
indukcji odpowiedzi immunologicznej (68). Dowiadcze-
kich komórek chorobowych i dlatego trudno porówny-
nia z komórkami linii embrionalnej (ang. embryonic stem
waæ badania na nich wykonywane. Efektywnoæ trans-
cells, ES) czy linii potworniakorakowej P19 (ang. terato-
fekcji pierwotnych linii komórkowych wynosi mniej ni¿
carcinoma cells) s¹ trudne ze wzglêdu na opornoæ na
kilka procent. Skutecznoæ zale¿y nie tylko od typu ko-
transfekcjê (68). siRNA charakteryzuje siê wysok¹ efek-
mórek, ale tak¿e od liczby pasa¿y i stopnia konfluencji
tywnoci¹ wprowadzenia do komórki oraz mo¿liwoci¹
komórek (45). Wiele linii komórkowych wykazuje tak¿e
osi¹gniêcia tam wysokiego stê¿enia. Zastosowanie takich
genetyczn¹ niestabilnoæ, co prowadzi do utraty klonal-
Tabela 5. Linie komórkowe wykorzystywane w badaniach
noci oraz zmian kariotypu i fizjologii (15). W zale¿noci
wyciszania genów przez siRNA (68, 63).
od podatnoci komórek na transfekcjê proponowane s¹
ró¿ne rozwi¹zania metodyczne (ryc. 1).
Wstêpne badania skutecznoci interferencji RNA
w uk³adach komórkowych pozwalaj¹ oceniæ stosowane
cz¹steczki siRNA lub shRNA. Zaproponowano dwufunk-
cjonaln¹ konstrukcjê siRNA-DNA zwan¹ crook siRNA
(34). Umo¿liwia ona wykrywanie bardzo niskiego pozio-
mu siRNA efektywnego dla RNAi, pobranej przez komórki
iloci siRNA, lokalizacji komórkowej, rozprowadzenia do
tkanek, a tak¿e farmakokinetyki. W crook siRNA do
koñca 3 sensownej nici siRNA do³¹czona jest struktura
DNA typu spinka do w³osów odporna na dzia³anie nu-
kleaz (34). Umo¿liwia to zarówno efektywne wyciszanie
genów jak i amplifikacjê PCR, przy minimalnej wykry-
Dla skutecznego zastosowania lub efektywnego bada-
nia procesu interferencji niezbêdny jest wybór odpowied-
niego celu molekularnego (mRNA) o znanej sekwencji. Jego
wyciszenie powinno daæ widoczny lub mo¿liwy do ob-
serwacji efekt. W niektórych dowiadczeniach in vitro
pokazano efekty uboczne zwi¹zane z wielokierunkowym
dzia³aniem siRNA. Dla ich unikniêcia dany gen nie powi-
Ludzkie komórki nab³onka ¿y³y pêpkowej
nien zawieraæ wielu sekwencji homologicznych. Celowe
jest skorzystanie z baz danych sekwencji genomowych
BLAST (NCBI Unigene, EST, Celera) dla weryfikacji popraw-
noci celu RNAi (47). Dostêpne s¹ algorytmy u³atwiaj¹ce
projektowanie skutecznych siRNA (tab. 6). Wprowadza-
j¹c tam numer dostêpu (ang. accession number) lub se-
kwencjê genu koduj¹cego cel terapeutyczny, otrzymuje-
my listê kompatybilnych oligonukleotydów. Wielu sku-
tecznych siRNA nie mo¿na jednak przewidzieæ tylko na
podstawie analizy in silico. Wszystkie mo¿liwe sekwen-
cje okrelonej d³ugoci powinny byæ analizowane w li-
niach komórkowych dla upewnienia, czy wszystkie siR-
NA zosta³y zidentyfikowane (67). Jest to bardzo kosztow-
ne. £atwiejsz¹ alternatywê stanowi stworzenie genowo-
specyficznej biblioteki siRNA poprzez przygotowanie
Tabela 6. Adresy internetowe wybranych algorytmów do projektowania siRNA.
http://katahdin.cshl.org:9331/RNAi/html/rnai.html
http://www.rnaiweb.com/RNAi/siRNA_Design/index.html
http://hydra1.wistar.upenn.edu/Projects/siRNA/siRNAindex.htm
https://www.genscript.com/ssl-bin/app/rnai?op=help
http://www.ambion.com/techlib/misc/siRNA_finder.html
http://i.cs.hku.hk/~sirna/software/sirna.php
http://www.rockefeller.edu/labheads/tuschl/sirna.html
https://rnaidesigner.invitrogen.com/rnaiexpress/
http://203.199.182.73/gnsmmg/databases/sirna/dstho.html
http://itb1.biologie.hu-berlin.de/~nebulus/sirna
mieszaniny cz¹steczek o mo¿liwej sekwencji, ekspresji
o d³ugoci poni¿ej 30 par zasad (60). W przypadku d³u¿-
w liniach komórkowych, selekcji wybranych fenotypów
szych cz¹steczek mo¿liwa jest aktywacja odpowiedzi
i na tej podstawie identyfikacji skutecznej sekwencji
interferonowej, prowadz¹cej do apoptozy. Nastêpuje
aktywacja szlaku Jak-Stat (ang. Janus kinase - signal trans-
ducers and activators of transcription) oraz wzrostu po-
ziomu ekspresji genów stymulowanych interferonem (in-
Warunkiem bezpiecznego wykorzystania RNAi w te-
terferon-stimulated genes, ISGs) (63). Transfekcja siRNA
rapii jest brak efektów ubocznych (ang. off-target effects).
swoistego dla GAPDH powodowa³a co najmniej dwu-
siRNA mo¿e funkcjonowaæ jako miRNA i na odwrót,
krotny wzrost ekspresji 52 genów ISG (63). W wiêkszoci
wykazuj¹c wieloraki wp³yw na ekspresjê genów (33).
przypadków poziom ekspresji zale¿ny by³ od iloci do-
Wykorzystanie ró¿nych siRNA dla jednego docelowego
starczonego siRNA, przy stê¿eniu 100 nM powoduj¹c
mRNA mo¿e powodowaæ uzyskanie ró¿nych niespecy-
ponad dziesiêciokrotny wzrost ekspresji. D³ugi RNA
ficznych wzorców ekspresji dla ka¿dej z u¿ytych cz¹ste-
o strukturze spinki do w³osów (ang. long-hairpin RNA,
czek. Mimo ¿e jedn¹ z cech RNAi jest specyficznoæ,
lhRNA), ulegaj¹cy ekspresji w komórkach ssaków z wek-
efekty off-target stanowi¹ potencjalny problem. Nadal
tora plazmidowego nie indukowa³ odpowiedzi immuno-
nieznane s¹ parametry okrelaj¹ce minimalny poziom
logicznej (37). Aktywacja uk³adu immunologicznego przez
homologii niezbêdny do efektywnego i bezpiecznego
siRNA i zwi¹zana z tym toksycznoæ s¹ zale¿ne od se-
wyciszania genów (69). Efektów ubocznych nie obser-
kwencji nukleotydowej (35). Zidentyfikowano immunosty-
wowano, gdy wprowadzano dsRNA do organizmów pry-
muluj¹ce motywy bogate GU, co pozwala na zaprojekto-
mitywnych. Wydaje siê, ¿e siRNA powstaj¹ce z dsRNA
wanie siRNA, który mo¿e wyciszaæ gen, jednoczenie in-
s¹ dok³adnie wybierane przez Drosha i Dicer oraz inne
dukuj¹c minimaln¹ odpowied interferonow¹ (35).
komponenty endogennej maszynerii RNAi, które mog¹
Przypuszcza siê, ¿e stymulacja odpowiedzi immuno-
mieæ aktywnoæ korekcyjn¹, która zabezpiecza przed
logicznej wywo³ana obecnoci¹ obcego RNA zachodzi
niespecyficznym wyciszaniem (14). siRNA powstaj¹cy
z udzia³em dwóch typów bia³ek: transmembranowych
z prekursora dsRNA w wyniku dzia³ania kompleksów en-
receptorów TLR (ang. Toll-like receptor) oraz cytoplazma-
zymatycznych w komórkach ssaków indukuje niespecy-
tycznych receptorów typu PKR (ang. protein kinase R) i pro-
ficzne efekty w znacznie mniejszym stopniu. Wi¹¿e siê
duktu genu RIG-1 (ang. retinoic acid-inducible gene 1),
to w tym przypadku z odpowiedzi¹ interferonow¹ (14).
(ryc. 2) (61). TLR znajduj¹ siê na b³onie komórkowej oraz
D³ugie dsRNA efektywnie wyciszaj¹ geny w komór-
w innych wewn¹trzkomórkowych kompartmentach.
kach owadów i komórkach embrionalnych ssaków, nie
W endosomach siRNA zawieraj¹cy motyw immunosty-
aktywuj¹c uk³adu immunologicznego (68). Do dojrza³ych
muluj¹cy aktywuje TLR3, TLR7, TLR8 i TLR9, co prowa-
komórek ssaków wprowadziæ mo¿na wy³¹cznie siRNA
dzi do uaktywnienia czynników transkrypcyjnych NF-ê ,
Rycina 2. Odpowied komórki na wprowadzenie ró¿nych typów dsRNA. A. Wektor wirusowy zawieraj¹cy sekwencjê shRNA inte-
gruje z genomem i ulega transkrypcji. Powsta³y shRNA jest transportowany do cytoplazmy przez eksportynê-5, hydrolizowany przez
Dicer do siRNA zawieraj¹cego niesparowane koñce i prowadzi do specyficznej degradacji docelowego mRNA. Odpowied immu-
nologiczna nie ma miejsca, gdy¿ zosta³y wykorzystane naturalne mechanizmy RNAi. B. Syntetyczny siRNA dostarczony do organi-
zmu w zawiesinie liposomalnej, dostaje siê do endosomu, gdzie aktywuje receptory TRL, co prowadzi do transdukcji sygna³u do j¹-
dra komórkowego i ekspresji genów cytokin zapalnych i interferonu. siRNA zawieraj¹cy dwa niesparowane nukleotydy przy koñcu
3 mo¿e wejæ na szlak RNAi, a siRNA ze sparowanymi koñcami aktywuje odpowied immunologiczn¹. C. Wprowadzony do ko-
mórki d³ugi dsRNA aktywuje kinazê bia³kow¹ PKR i RNazê L, co prowadzi do degradacji RNA i zablokowania inicjacji translacji,
a w konsekwencji do apoptozy. TLR ang. Toll-like receptor, PKR kinaza bia³kowa zale¿na od dsRNA, RIG-1 ang. retinoic acid-
inducible gene I, EIF2 podjednostka czynnika inicjacji translacji.
IRF-3 oraz IRF-7, które stymuluj¹ ekspresjê genów IFN.
prowadzi do ca³kowitego zablokowania syntezy bia³-
Helikazowa domena bia³ka RIG-1 w cytoplazmie rozpo-
ka. Komórka kierowana jest na drogê apoptozy. Wi¹-
znaje dsRNA, a pozosta³e domeny aktywuj¹ szlak pro-
zanie dsRNA przez syntazê 2',5'-oligoA powoduje jej
wadz¹cy do syntezy interferonu i cytokin zapalnych (61).
aktywacjê i syntezê oligonukleotydów adenylowych
siRNA zawieraj¹cy dwa niesparowane nukleotydy przy
(pppA(2p5A) ), które aktywuj¹ RNazê L, powoduj¹c¹
koñcu 3 nie aktywuje bia³ka RIG-1, w odró¿nieniu od
degradacjê RNA (45). Chemiczne cechy dsRNA, poziom
siRNA nie zawieraj¹cego niesparowanych koñców (44).
ekspresji lub dawki oraz sposób dostarczenia RNA do
Tak¿e shRNA ulegaj¹ce ekspresji z wektora nie aktywuj¹
komórki mog¹ wp³ywaæ na poziom aktywacji odpowie-
odpowiedzi interferonowej. Natomiast reakcjê tê stymu-
luje syntetyczny siRNA dostarczony za pomoc¹ nonika
Istnieje mo¿liwoæ wspó³zawodnictwa egzogennych
lipidowego (55). Modyfikacja chemiczna siRNA mo¿e
siRNA z endogennymi miRNA o dostêp do RISC (ang.
utrudniæ rozpoznanie przez TLR, co u³atwi³oby stosowa-
RNA-induced silencing complex). miRNA odgrywaj¹ rolê
nie siRNA bez wzbudzania odpowiedzi immunologicz-
regulatorów ekspresji genów. Zaburzenie ich funkcjono-
wania mo¿e powodowaæ komplikacje terapeutyczne.
Aktywacja kinazy bia³kowej PKR nastêpuje po zwi¹-
W niektórych przypadkach nadekspresja genów miR
zaniu dsRNA. Hamuje ona translacjê na skutek fosforyla-
mo¿e prowadziæ do onkogenezy, z kolei w komórkach
cji podjednostki czynnika inicjacji translacji EIF2 , co
nowotworowych obserwowano zredukowany poziom
niektórych miRNA (69). Obni¿enie iloci stosowanego
przez Dicer oraz efektywniejszym wprowadzeniem do
siRNA mo¿e przyczyniæ siê do obni¿enia ryzyka wyst¹-
RISC (2). Istnieje zale¿noæ skutecznoci disRNA od wy-
pienia efektów niespecyficznych (28). siRNA wykazuj¹-
ciszanego genu. dsRNA o d³ugoci oko³o 200 nukleoty-
cy pe³n¹ komplementarnoæ do docelowego genu mo¿e
dów skutecznie wycisza³ gen tenascyny-C (TN-C) in vitro
powodowaæ, oprócz degradacji w³aciwego mRNA, tak-
i in vivo (84). Strategiê oparto na za³o¿eniu, ¿e Dicer wi¹¿e
¿e spadek poziomu bia³ek w komórce (18). Zaprojekto-
i hydrolizuje dsRNA do dupleksów o d³ugoci 21-25 nu-
wany siRNA, zawieraj¹cy trzy do czterech nukleotydów
kleotydów, które po wprowadzeniu do RISC mog¹ zwie-
niesparowanych z nukleotydami transkryptu docelowe-
lokrotniæ efekt wyciszenia, poprzez wi¹zanie siê do wie-
go inhibitora 1A kinazy zale¿nej od cyklin (ang. cyclin-
lu miejsc mRNA wiêkszej iloci cz¹steczek (84). Na po-
dependent kinase inhibitor 1A, CDKN1A), wp³ywa³ g³ów-
dobnej zasadzie projektowano d³ugi RNA o strukturze
nie na poziom bia³ek, w sposób podobny do miRNA (33).
spinki do w³osów (long-hairpin RNA, lhRNA) w celu wy-
Badano wp³yw d³ugotrwa³ego wysokiego poziomu
ciszenia genów wirusa HIV (37). Ekspresja lhRNA pro-
ekspresji shRNA na w¹troby doros³ych myszy zbadano
wadzi do powstania siRNA skierowanych przeciwko wielu
stosuj¹c 49 ró¿nych wektorów AAV/shRNA, unikalnych
regionom genu nef, co powodowa³o wydajniejsz¹ inhi-
pod wzglêdem d³ugoci i sekwencji przeciwko szeciu
genom docelowym (24). 36 sporód nich powodowa³o
W wektorach ekspresyjnych siRNA wykorzystywane
powa¿ne uszkodzenia w¹troby zale¿ne od stosowanej
s¹ g³ównie promotory polimerazy RNA III (Pol III) oraz II
dawki, z czego 23 ostatecznie prowadzi³y do mierci
(Pol II). Ze wzglêdu na obfitoæ komórkowych niekodu-
ponad 150 zwierz¹t w ci¹gu 2 miesiêcy (12, 24). Zacho-
j¹cych RNA transkryptów Pol III, czêsto wykorzystuje
rowalnoæ by³a zwi¹zana z obni¿eniem iloci endogen-
siê promotory dla tej polimerazy w badaniach nad RNAi
nych miRNA, wskazuj¹c na prawdopodobn¹ konkuren-
w komórkach ssaków. Promotory te umo¿liwiaj¹ produk-
cjê pomiêdzy miRNA i egzogennymi shRNA o ograni-
cjê cz¹steczek RNA o cechach charakterystycznych dla
czon¹ iloæ czynników komórkowych zaanga¿owanych
siRNA nie zawieraj¹cych struktury cap przy koñcu 5'
w funkcjonowanie RNAi (24). Elementem ograniczaj¹-
oraz poliadenylowanego koñca 3'. Wród promotorów
cym funkcjonowanie miRNA jest eksportyna-5, ulegaj¹-
Pol III do produkcji siRNA in vivo wykorzystuje siê pro-
motor U6 genu ma³ych j¹drowych RNA (ang. small nuc-
lear RNA, snRNA) oraz promotor H1 genu RNazy P (77).
Wprowadzanie cz¹steczek siRNA do komórek
W przypadku promotora H1, istotn¹ zalet¹ jest uzyskiwa-
siRNA mo¿na otrzymaæ in vitro, metod¹ syntezy che-
nie transkryptów zawieraj¹cych niesparowane koñce 3'.
micznej, transkrypcji in vitro oraz poprzez trawienie d³u-
Wykorzystywane s¹ równie¿ promotory typu II. W kilku
gich dsRNA enzymami RNazaIII/Dicer. Mo¿liwa jest tak-
przypadkach okaza³o siê, ¿e s¹ one efektywniejsze od pro-
¿e ich ekspresja in vivo. Regulacja ekspresji genów za
motorów typu III oraz prowadz¹ do zagêszczenia siRNA
pomoc¹ siRNA jest przemijaj¹ca i czêsto trwa przez 3-5
w cytoplazmie. Cz¹steczki ulegaj¹ce ekspresji z wykorzy-
staniem promotorów U6 wystêpuj¹ g³ównie w j¹drze ko-
Dla wyciszania genów w komórkach embrionalnych
mórkowym. Dicer wykazuje wy¿sz¹ aktywnoæ w cyto-
Drosophila melanogaster najefektywniejsz¹ cz¹steczk¹ wy-
plazmie i przetwarzanie shRNA w aktywny siRNA mo¿e
ciszaj¹c¹ jest siRNA o d³ugoci 21 nukleotydów i dwoma
niesparowanymi nukleotydami przy koñcu 3 (16). Dalsze
Wykorzystanie promotorów Pol II poza pewnymi ty-
badania wykaza³y, ¿e skutecznoæ cz¹steczek siRNA wzra-
pami komórek jest jednak bardzo ograniczone. Pol II
sta wraz z d³ugoci¹ dupleksu a¿ do 27pz (36). D³u¿sze
uczestniczy w transkrypcji genów koduj¹cych bia³ka.
cz¹steczki charakteryzuje mniejsza skutecznoæ (2).
Transkrypty maj¹ typowe modyfikacje koñców 3' i 5', ta-
W mechanizmie interferencji RNA, dsRNA jest roz-
kie jak struktura cap czy poli(A) Nie odpowiada to struk-
poznawany i hydrolizowany przez Dicer w odpowied-
turalnym wymaganiom siRNA. Z wyj¹tkiem kilku typów
niej odleg³oci od koñca helikalnego (42). Mo¿na wyko-
komórek d³ugie dsRNA uzyskane dziêki promotorom Pol
rzystaæ Dicer do wyboru i wyciêcia najbardziej efektyw-
II, s¹ natychmiast transportowane do cytoplazmy, gdzie
nej cz¹steczki siRNA z wprowadzonej do komórki nici
indukuj¹ odpowied interferonow¹ (77). Uda³o siê uzy-
shRNA lub dsRNA. Istnieje mo¿liwoæ zaprojektowania
skaæ efekt wyciszenia z u¿yciem wektorów, zawieraj¹-
disRNA (ang. Dicersubstrate siRNA), który po hydroli-
zie przez Dicer dawa³by specyficzne siRNA o d³ugoci
Ekspresja dwuniciowych RNA w komórkach mo¿liwa
21 nukleotydów. Uzyskane cz¹steczki disRNA powodo-
jest na dwóch drogach. Pierwsza oparta jest o syntezê
wa³y maksymaln¹ inhibicjê ekspresji genów przy ni¿szych
dwóch komplementarnych nici siRNA o d³ugoci 21-23
stê¿eniach, a wyciszenie trwa³o d³u¿ej (2). Spowodowa-
nukleotydów z dwóch ró¿nych promotorów U6, z kaset
ne jest to prawdopodobnie rozpoznaniem i hydroliz¹
zlokalizowanych tandemowo na tym samym wektorze
Tabela 7. Mechanizmy indukcji ekspresji shRNA. DSE - ang. distal sequence element; Dox - doksycyklina; GAL4 - N-
koñcowy fragment transkrypcyjnego aktywatora dro¿d¿y; KRAB - rodzina transkrypcyjnych represorów zwi¹zanych
z Kruppel box; PSE - ang. proximal sequence element; RXR - receptor Retinoidu X; tetO - operator tetracyklinowy; tTR
- bia³ko represorowe tetracykliny; VgEcR - zmodyfikowany receptor ekdyzonu, neo - neomycyna.
Sekwencja tetO zlokalizowana bezporednio poni¿ej TATA box.
W nieobecnoci Dox, tTR wi¹¿e siê do tetO i hamuje transkrypcjê shRNA.
Dodanie Dox powoduje dysocjacjê tTR, co prowadzi do aktywacji jednostki
Wiele sekwencji tetO znajduje siê pomiêdzy elementami promotora
Pol III DSE i PSE oraz TATA box. Taka budowa poprawia kooperatywne
wi¹zanie tTR do sekwencji tetO i jednoczenie blokuje komunikacjê pomiêdzy
ka¿dym z tych elementów. Dox powoduje dysocjacjê tTR, co prowadzi
do aktywacji jednostki transkrypcyjnej i ekspresji shRNA
U¿ycie bia³ka hybrydowego tTR-KRAB, kontrolowanego tetracyklin¹.
W nieobecnoci Dox, tTR-Krab wi¹¿e siê do tetO i hamuje transkrypcjê
shRNA. Dodanie Dox powoduje dysocjacjê tTR-KRAB, co prowadzi
do aktywacji jednostki transkrypcyjnej i ekspresji shRNA
Obejmuje trzy wektory retrowirusowe: ekspresja dwóch z nich prowadzi
do syntezy dwóch czynników transkrypcyjnych VgEcR i RXR, a trzeciego
do syntezy produktu genu po³¹czonego Gal4, bêd¹cego porednim
aktywatorem, pozostaj¹cego pod kontrol¹ indukowalnego promotora.
Ekdyzon powoduje dimeryzacjê czynników transkrypcyjnych i wi¹zanie
siê ich do indukowalnego promotora, co aktywuje aktywator poredni
GAL4. £¹czy siê on do czterech miejsc wi¹zania GAL4 w DNA,
aktywuj¹c promotor U6, co prowadzi do aktywacji jednostki
U myszy transgeniczej nie dochodzi do ekspresji U6-ploxPneo-Fgfr2
i wyciszenia genu Fgfr2. Promotor U6 jest nieaktywny z powodu insercji
sekwencji neo. Po skrzy¿owaniu z mysz¹, u której w linii komórek
rozrodczych dochodzi do ekspresji Cre, dochodzi do wyciêcia neo
Promotor H1 ma wprowadzony operator lac. Zastosowanie w transgenicznych
liniach komórkowych, w których ekspresji ulega represor lac.
Po zastosowaniu IPTG, represor lac oddysocjowuje i mo¿liwa jest transkrypcja.
lub na dwóch ró¿nych plazmidach (77). Drugi sposób
¿e tRNA-shRNA, ulegaj¹cy ekspresji z wykorzystaniem
wymaga u¿ycia jednego promotora, aby uzyskaæ ekspre-
promotora ludzkiego genu dla tRNAVal prowadzi do spe-
sjê siRNA o strukturze spinki do w³osów (shRNA), przy
cyficznej inhibicji ekspresji genu fuzyjnego PML-RAR
czym koniec 3' 19- do 21-nukleotydowej sekwencji po-
zarówno in vitro jak i in vivo (49).
³¹czony jest z koñcem 5' komplementarnej sekwencji tej
Badania wyciszania genów zwi¹zanych z cyklem ko-
samej d³ugoci poprzez pêtlê zawieraj¹c¹ 3 do 9 nukle-
mórkowym, apoptoz¹ i rozwojem s¹ bardzo trudne. Hy-
otydów (77). W obu przypadkach powstaj¹ dwuniciowe
droliza mRNA krytycznego dla organizmu na danym eta-
cz¹steczki siRNA, przy czym drugi sposób wymaga za-
pie rozwoju mo¿e prowadziæ do zaburzeñ i uniemo¿liwiæ
anga¿owania przez komórkê enzymu Dicer.
uzyskanie wyników na kolejnych etapach. Aby temu za-
Istnieje mo¿liwoæ uzyskania ekspresji shRNA zwi¹-
pobiec mo¿na stosowaæ wektory, zawieraj¹ce promotory
zanych z tRNA przy koñcu 3, co zwiêksza efektywnoæ
warunkowe (ang. conditional promoters) (tab. 7, ryc. 3).
dostarczenia cz¹steczek do cytoplazmy (2). Wykazano,
W tym celu wykorzystano promotory Pol III, pomimo ich
nie wektorów wirusowych daje mo¿liwoæ uzyskania sta-
bilnego wyciszenia docelowego genu, w zwi¹zku z mo¿-
liwoci¹ integracji genomu wirusa z genomem gospoda-
rza i d³ugotrwa³ej transkrypcji wyciszaj¹cego RNA. Wek-
tory wirusowe ró¿ni¹ siê miêdzy sob¹ typem preferowa-
nych komórek docelowych, ³atwoci¹ zastosowania in
Rycina 3. Zale¿noæ syntezy siRNA od tetracykliny (Tet) lub do-
vitro i in vivo, zdolnoci¹ do integracji z genomem go-
ksycykliny (Dox). TetR represor tetracyklinowy; TetO opera-
spodarza lub jej brakiem, poziomem immunogennoci
oraz maksymalnym mo¿liwym rozmiarem insertu.
mniejszej wszechstronnoci w porównaniu z Pol II. Tego
Kontrola procesu interferencji RNA mo¿e byæ zreali-
typu konstrukcje cile reguluj¹ transkrypcjê siRNA w ko-
zowana w przypadku po³¹czenia cech wektorów wiru-
mórkach, w których zachodzi przejciowa lub stabilna
sowych i systemów ekspresji siRNA opartych na promo-
ekspresja represora Tet (TetR) (77). Sekwencja operatora
torach warunkowych. W przeprowadzonych dowiadcze-
tertracyklinowego TetO umieszczona jest powy¿ej se-
niach in vitro, równoczesna infekcja wektorem adenowi-
kwencji promotorów H1, U6 lub 7SK. Przy³¹czenie siê
rusowym, zawieraj¹cym system ekspresji siRNA pozosta-
represora do operatora hamuje transkrypcjê. Tetracykli-
j¹cy pod kontrol¹ promotora H1, zale¿nego od doksycy-
na lub doksycyklina powoduj¹ dysocjacjê kompleksu, co
kliny oraz wektorem adenowirusowym koduj¹cym repre-
prowadzi do aktywacji jednostki transkrypcyjnej i eks-
sor tetracyklinowy, doprowadzi³a do inhibicji ekspresji
presji interferuj¹cego RNA. Wektory tego typu umo¿li-
genów p53 i c-Myc w sposób zale¿ny od doksycykliny
wiaj¹ kontrolê poziomu docelowego bia³ka poprzez re-
i dawki wirusa (30). W ten sposób uda³o siê uzyskaæ po-
gulacjê stê¿enia czynnika indukcyjnego. Trwaj¹ prace nad
wykorzystaniem wektorów, w których ekspresja interfe-
dsRNA mo¿e zostaæ wprowadzony do komórki na
rencyjnego RNA zale¿na jest od dwóch czynników kon-
drodze elektroporacji, mikroiniekcji, zanurzania organi-
trolnych ekdyzonu oraz produktu genu Gal4 (25). Pro-
zmu w roztworze dsRNA, drog¹ pokarmow¹ lub z wyko-
motory warunkowe Pol III umo¿liwiaj¹ regulacjê ekspre-
rzystaniem wektorów plazmidowych lub wirusowych.
sji shRNA tkankowo- lub komórkowospecyficznej po-
Metody te by³y stosowane w stosunku do ró¿nych orga-
przez ekspresjê aktywatora GAL4 pozostaj¹cego pod kon-
nizmów (1). Podanie myszy o wadze 20 g zastrzyku roz-
trol¹ tkankowospecyficznych promotorów (47).
tworu siRNA o objêtoci 1ml odpowiada iniekcji 3,5 l
Wektory maj¹ce promotory warunkowe uda³o siê za-
roztworu pacjentowi o wadze 70 kg (5). Istnieje mo¿li-
stosowaæ w dowiadczeniach in vitro. Pokazano kontro-
woæ wprowadzenia siRNA do organizmu drog¹ inhala-
lowane wy³¹czenie genu DMNT1 (DNA metylotransfe-
cji. Wprowadzony w ten sposób siRNA u myszy zahamo-
raza-1) w ludzkich komórkach rakowych, prowadz¹ce
wa³ infekcjê wywo³an¹ wirusami RSV (ang. respiratory syn-
do zahamowania ich wzrostu, wyciszenia genu CXCR4
cytial virus) oraz PIV (ang. parainfluenza virus) (4, 79). Dla
(receptor CXC4, zwi¹zany z ruchliwoci¹) w komórkach
lokalnego dostarczania cz¹steczek RNAi istotny jest wy-
raka piersi, znacznego zahamowania ich przemieszcza-
bór powierzchni wnikania terapeutyku. B³ony luzowe
nia; oraz do wy³¹czenia genu -kateniny w komórkach
s¹ miejscem wnikania wielu czynników infekcyjnych,
raka okrê¿nicy, uzyskuj¹c zahamowanie cyklu komór-
dlatego wydaj¹ siê byæ doskona³ym celem dla RNAi (59).
kowego i wzrostu (47). Opisany system indukcyjny zo-
Istnieje szansa na stworzenie leku opartego na technolo-
sta³ tak¿e sprawdzony in vivo u myszy, u której wystêpo-
gii RNAi bez koniecznoci wykonywania zastrzyków, ale
wa³y przerzuty nowotworu, przy czym ekspresja siRNA
poprzez kontakt z b³on¹ luzow¹ dróg oddechowych,
nie podlega³a w tym przypadku tak cis³ej regulacji jak
rozrodczych, czy uk³adu pokarmowego.
w przypadku badañ in vitro na wyselekcjonowanych ko-
W wiêkszoci przypadków plazmidy ulegaj¹ przejcio-
Badania wyciszania genów RNAi wkroczy³y na etap
wej ekspresji z wytworzeniem siRNA. Drugi typ noni-
badañ klinicznych. W fazie I eksperymentalny lek jest
ków wykorzystywanych do uzyskania wyciszenia doce-
testowany po raz pierwszy na ma³ej grupie ludzi (20-80
lowych genów stanowi¹ wektory wirusowe. W celu do-
osób), w celu okrelenia bezpiecznej dawki leku oraz
starczenia do komórek ekspresyjnych kaset siRNA mo¿na
identyfikacji efektów ubocznych. W fazie II badaniu pod-
wykorzystaæ trzy rodziny cz¹steczek wirusowych: oparte
lega wiêksza liczba osób stosuj¹cych dany lek (100-300).
na retrowirusach (zarówno onkowirusy jak i lentiwirusy),
Celem tego etapu jest sprawdzenie efektywnoci prepa-
adenowirusach oraz wirusach AAV (ang. adeno-associa-
ratu oraz dalsze sprawdzanie jego bezpieczeñstwa. Eks-
ted viruses) (77), które do replikacji potrzebuj¹ wirusa wspo-
perymentalny rodek podawany wiêkszej grupie ludzi
magaj¹cego (adenowirusa lub herpeswirusa). Zastosowa-
(1000-3000) charakteryzuje fazê III badañ klinicznych.
Tabela 8. Badania kliniczne RNAi (5, 17). AMD ang Age-related macular degeneration, degeneracja plamki ¿ó³tej,
DMO Diabetic macular oedema, cukrzycowy obrzêk plamki ¿ó³tej, IND proba do FDA o pozwolenie na testowa-
Jej celem jest potwierdzenie jego efektywnoci, monito-
age-related macular degeneration, AMD) poprzez inhi-
rowanie efektów ubocznych, porównanie z powszech-
bicjê ekspresji genu VEGF (ang. vascular endothelial
nie stosowanymi lekami oraz zbieranie informacji umo¿-
growth factor). Obecnie w stanie nieuleczalnym AMD
liwiaj¹cych w przysz³oci bezpieczne jego stosowanie.
jest 1.65 milionów Amerykanów (57). Acuity Pharmaceu-
W fazie IV zbierane s¹ dodatkowe informacje, ju¿ po
ticals sugeruje, ¿e na wiecie w roku 2013 na AMD bêdzie
wprowadzeniu leku na rynek. Badania te dotycz¹ ryzyka
chorowaæ 11 milionów ludzi (57). Badania przeprowadzo-
stosowania leku, korzyci oraz dawki optymalnej (75).
ne na grupie 129 pacjentów wskazuj¹, ¿e wstrzykniêty do
Chorobotwórczy gen ApoB (apoliproteina B) zosta³
oka Bevasiranib redukuje wzrost naczyñ krwiononych,
wyciszony u ma³p poprzez wprowadzenie siRNA do
co nieznacznie poprawia wzrok (57). Przy niskich daw-
krwiobiegu (9). siRNA specyficzny wobec ApoB w posta-
kach efekt utrzymywa³ siê przez kilka miesiêcy, przy wy-
ci cz¹stek SNALP (ang. stable nucleic acid lipid particles)
¿szych o wiele d³u¿ej. Nie obserwowano szkodliwych efek-
podany w wysokiej dawce (1 i 2.5 mg kg1) akumulowa³
tów ubocznych z wyj¹tkiem przewidywanej opuchlizny
siê w w¹trobie po 48 godzinach powoduj¹c wyciszenie
i stanu zapalnego w miejscu wstrzykniêcia leku (57). Nie-
ponad 90% (83). Efekt utrzyma³ siê przez 11 dni, nie za-
wykluczone, ¿e Bevasiranib jak i inne podobne terapeuty-
obserwowano efektów toksycznych dla organizmu (9).
ki bêd¹ stosowane ³¹cznie z lekami przeciwnowotworo-
Wprowadzenie systemiczne siRNA do zwierz¹t naczel-
wymi, które tak¿e blokuj¹ efekty VEGF.
nych okaza³o siê skuteczne i bezpieczne, co daje nadziejê
Pierwsze wyniki badañ klinicznych wydawa³y siê
zgodne z teori¹. Pod wp³ywem leku dochodzi³o do efek-
Firmy biotechnologiczne rozpoczê³y lub planuj¹ roz-
tywnej redukcji rozmiarów nowotworów oraz iloci cz¹-
poczêcie badañ klinicznych leków opartych na techno-
stek wirusowych u pacjentów (56). Dok³adna analiza
logii RNAi (tab. 8). Badania dotycz¹ terapii chorób wiru-
wyników wykazuje, ¿e uzyskane rezultaty spowodowa-
sowych jak i wieloczynnikowych. Dotychczas prace ba-
ne by³y raczej wzrostem produkcji interferonu przez uk³ad
dawcze obejmuj¹ pierwsz¹ fazê badañ klinicznych. Kil-
immunologiczny w odpowiedzi na obcy RNA, ni¿ spe-
ka firm skupia siê na terapii degeneracji plamki ¿ó³tej (ang.
cyficznym wyciszeniem docelowych genów (56).
rynku odzwierciedlaj¹ czas zaadaptowania danej techni-
Od czasu opisania istoty RNAi pod koniec ubieg³ego
ki. Pierwsz¹ opracowan¹ metod¹ by³y oligonukleotydy
wieku, technologia ta zosta³a szybko zaadaptowana w la-
RNA, nastêpnie techniki oparte o wektory. Wzglêdnie nowy
boratoriach badawczych. Interferencja RNA w odró¿nie-
obszar stanowi lecznictwo, obejmuj¹ce zastosowanie RNAi
niu od innych metod terapeutycznych wykorzystuje na-
do odkrywania nowych leków. Przewiduje siê, ¿e dochód
turalne elementy maszynerii komórkowej. siRNA w swym
zwi¹zany z technologi¹ RNAi bêdzie systematycznie wzra-
dzia³aniu przypominaj¹ katalizatory jedna cz¹steczka
sta³, osi¹gaj¹c w 2008 roku 185 milionów dolarów.
mo¿e wyciszyæ wiele cz¹steczek mRNA. Zapewnia to
Szacuje siê, ¿e technologia RNAi obejmie oko³o 10%
rynku leków, co oznacza miliardy dolarów zysków. Po-
Od 1998 roku roczna liczba publikacji na temat RNAi
mimo tego, ¿e zastosowanie RNAi budzi nadzieje w kil-
wzros³a z kilkunastu do kilkuset (31). Aby RNAi by³o sku-
ku przypadkach, wielu obserwatorów twierdzi, ¿e nie
teczn¹ metod¹ terapeutyczn¹, nale¿y przezwyciê¿yæ trud-
przejd¹ one pomylnie prób klinicznych i nie zostan¹
noci natury technicznej oraz finansowej, co oznacza
zatwierdzone w USA przez FDA przed rokiem 2008 lub,
produkcjê leku przy racjonalnych kosztach (56).
co bardziej prawdopodobne, przed 2013 (31).
Interferencja RNA nie zosta³a jeszcze wykorzystana jako
Odkrycie mo¿liwoci zastosowania interferencji RNA
metoda terapeutyczna, ale ju¿ teraz technologie RNAi przy-
jako technologii terapeutycznej silnie zaanga¿owa³o prze-
nosz¹ du¿e zyski. Rynek produktów opartych na RNAi
mys³. W badania RNAi w³¹czy³o siê ju¿ ponad 100 firm
w tym siRNA, oligonukleotydy RNA i wektory DNA, ko-
(66). Czêæ z nich dostarcza odczynniki i rozwi¹zania
duj¹ce siRNA przyniós³ dochód w roku 2003 w wysoko-
metodyczne niezbêdne do wykonania dowiadczeñ, resz-
ci 38 milionów dolarów (dane wed³ug Front Line Strate-
ta to firmy biotechnologiczne lub farmaceutyczne poszu-
gic Consulting, San Meteo, USA) (31) (ryc. 4). Rozmiary
kuj¹ce komercyjnych zastosowañ RNAi (tab. 9).
Tabela 9. Firmy biotechnologiczne wykorzystuj¹ce technologiê RNAi (31).
Michael Tolentino i Samuel RNAi przeciwko
przesiewowych genomu czeniu genu i u¿yciu
z u¿yciem RNAi w liniach ma³ych inhibitorów
Zastrze¿ona tajemnicza Leki przeciwko
8. Chen Y., Stamatoyannopoulos G., Song C.Z.: Downregulation of
CXCR4 by inducible small interfering RNA inhibits breast cancer
cell invasion in vitro. Cancer Res. 2003, 63, 4801-4804
9. Choi C.: RNAi works in monkeys. 2006, www.the-scientist.com/
10.Coburn G.A., Cullen B.R.: Potent and specific inhibition of human
immunodeficiency virus type 1 replication by RNA interference. J. Vi-
Rycina 4. Obszary rynku zwi¹zane z technologi¹ RNAi (31).
11.Coumoul X., Shukla V., Li C., Wang R.H., Deng C.X.: Conditional
Z mo¿liwych do wyró¿nienia trzech typów technik
knockdown of Fgfr2 in mice using Cre-LoxP induced RNA interfe-
anty-mRNA, obejmuj¹cych jednoniciowe antysensowne
oligonukleotydy, rybozymy i interferencjê RNA, ta ostat-
12.Couzin J.: RNAi safety comes under scrutiny, Science, 2006, 312, 1121
nia wydaje siê mieæ najlepsze perspektywy. O nieop³a-
13.Czauderna F., Santel A., Hinz M., Fechtner M., Durieux B., Fisch G.,
Leenders F., Arnold W., Giese K., Klippel A., Kaufmann J.: Inducible
calnoci technologii opartej na rybozymach mo¿e wiad-
shRNA expression for application in a prostate cancer mouse mo-
czyæ firma Sirna Therapeutics, która powsta³a z Ribozy-
me Pharmaceuticals, po tym jak kilka zaprojektowanych
14.Dillon C.P., Sandy P., Nencioni A., Kissler S., Rubinson D.A., Van
leków nie spe³nia³o oczekiwañ. Przemiana firmy na skra-
Parijs L.: RNAi as an experimental and therapeutic tool to study and
ju bankructwa, posiadaj¹cej jedynie 2 miliony dolarów
regulate physiological and disease processes. Annu. Rev. Physiol.
w gotówce, przynios³a w ci¹gu 18 miesiêcy a¿ 72 milio-
15.Echeverri C.J., Perrimon N.: High-throughput RNAi screening in
ny dolarów od nowych inwestorów. Nawet je¿eli tylko
cultured cells: a users guide. Nat. Rev. Genet. 2006, 7, 373-384
niewielka czêæ projektów opartych na RNAi znajdzie
16.Elbashir S.M., Martinez J., Patkaniowska A., Lendeckel W., Tuschl T.:
terapeutyczne zastosowanie, technologia ta mo¿e pro-
Functional anatomy of siRNAs for mediating efficient RNAi in Droso-
wadziæ do miliardów dolarów zysku z nowych leków (65).
phila melanogaster embryo lysate. EMBO J. 2001, 20, 6877-6888
17.Frantz S.: Safety concerns raised over RNA interference. Nat. Rev.
1. Agrawal N., Dasaradhi P.V.N., Mohmmed A., Malhotra P., Bhatna-
18.Gabryelska M., Nowak S., ¯ukiel R., Barciszewski J.: Interferencja RNA
gar R.K., Mukherjee S.K.: RNA interference: biology, mechanizm,
nowa technologia w leczeniu nowotworów. Neuroskop 2005, 7, 27-38
and applications. Microbiol. Mol. Biol. Rev. 2003, 67, 657-685
19.Gabryelska M., Nowak S., ¯ukiel R., Barciszewski J.: Uniwersalna
2. Amarzguioui M., Rossi J.J., Kim D.: Approaches for chemically syn-
metoda inhibicji ekspresji genów interferencja RNA. Na pograni-
thesized siRNA and vector-mediated RNAi. FEBS Lett. 2005, 579,
czu chemii i biologii 2005, Tom XII, 235-267
20.Gaur R.K.: RNA interference: a potential therapeutic tool for silencing spli-
3. B¹k D.: RNAi interferencja RNA skuteczny sposób na ciszê. Po-
ce isoforms linked to human diseases. Biotechniques, 2006, 40, 15-22
21.Ge Q., Filip L., Bai A., Nguyen T., Eisen H.N., Chen J.: Inhibition of
4. Bitko V., Musiyenko A., Shulyayeva O., Barik S.: Inhibition of respi-
influenza virus production in virus infected mice by RNA interfe-
ratory viruses by nasally administered siRNA, Nat. Med. 2005, 11,
rence. Proc. Natl. Acad. Sci. USA 2004, 101, 8676-8681
22.Glunde K., Raman V., Mori N., Bhujwalla Z.M.: RNA interference-
5. Cejka D., Losert D., Wacheck V.: Short interfering RNA (siRNA): tool
mediated choline kinase suppression in breast cancer cells induces
or therapeutic? Clin. Sci. 2006, 110, 47-58
differentiation and reduces proliferation. Cancer Res. 2005, 65,
6. Chang Y., Chang S.S., Lee H.H., Doong S.L., Takada K., Tsai C.H.:
Inhibition of the Epstein-Barr virus lytic cycle by Zta-targeted RNA
23.Gomez-Valades A.G., Vidal-Alabro A., Molas M., Boada J., Bermu-
interference. J. Gen. Virol. 2004, 85, 1371-1379
dez J., Bartrons R., Perales J.C.: Overcoming diabetes-induced hy-
7. Charames G.S., Bapat B.: Cyclooxygenase-2 knockdown by RNA
perglycemia through inhibition of hepatic phosphoenolpyruvate
interference in colon cancer. Int. J. Oncol. 2006, 28, 543-549
carboxykinase (GTP) with RNAi. Mol. Ther. 2006, 13, 401-10
24.Grimm D., Streetz K.L., Jopling C.L., Storm T.A., Pandey K., Davis
44.Marques J.T., Devosse T., Wang D., Zamanian-Daryoush M., Serbi-
C.R., Marion P., Salazar F., Kay M.A.: Fatality in mice due to oversa-
nowski P., Hartmann R., Fujita T., Behlke M.A., Williams B.R.:
turation of cellular microRNA/short hairpin RNA pathways. Nature
A structural basis for discriminating between self and nonself do-
uble-stranded RNAs in mammalian cells. Nat. Biotechnol. 2006, 24,
25.Gupta S., Schoer R.A., Egan J.E., Hannon G.J., Mittal V.: Inducible,
reversible, and stable RNA interference in mammalian cells. Proc.
45.McManus M.T., Sharp P.A.: Gene silencing in mammals by small
Natl. Acad. Sci. USA 2004, 101, 1927-1932
interfering RNAs. Nat. Rev. Genet. 2002, 3, 737-747
26.Hannon G.J., Rossi J.J.: Unlocking the potential of the human geno-
46.McRobert L., McConkey G.A.: RNA interference (RNAi) inhibits
me with RNA interference. Nature 2004, 431, 371-378
growth of Plasmodium falciparum. Mol. Biochem. Parasitol. 2002,
27.Harper S.Q., Staber P.D., He X., Eliason S.L., Martins I.H., Mao Q.,
Yang L., Kotin R.M., Paulson H.L., Davidson B.L.: RNA interference
47.Mittal V.: Improving the efficiency of RNA interference in mammals,
improves motor and neuropathological abnormalities in a Hunting-
tons disease mouse model. Proc. Natl. Acad. Sci. USA 2005, 102,
48.Mohmmed A., Dasaradhi P.V., Bhatnagar R.K., Chauhan V.S., Malho-
tra P.: In vivo gene silencing in Plasmodium berghei-a mouse malaria
28.Heidenreich O.: Oncogene suppression by small interfering RNAs.
model. Biochem. Biophys. Res. Commun. 2003, 309, 506-511
Curr. Pharm. Biotechnol. 2004, 5, 349-354
49.Oshima K., Kawasaki H., Soda Y., Tani K., Asano S., Taira K.: Maxi-
29.Higuchi M., Tsutsumi R., Higashi H., Hatekeyama M.: Conditional
gene silencing utilizing the lac repressor reveals a role of SHP-2 in
zymes and small hairpin-type RNAs that are driven by a tRNA pro-
cagA-positive Helicobacter pylori pathogenicity. Cancer Sci. 2004,
moter specifically cleave a chimeric gene associated with leukemia
in vitro and in vivo. Cancer Res. 2003, 63, 6809-6814
30.Hosono T., Mizuguchi H., Katayama K., Xu Z., Sakurai F., Ishii-Wa-
50.Pai S.I., Lin Y.Y., Macaes B., Meneshian A., Hung C.F., Wu T.C.:
tabe A., Kawabata K., Yamaguchi T., Nakagawa S., Mayumi T., Hay-
Prospects of RNA interference therapy for cancer. Gene. Ther. 2006,
akawa T.: Adenovirus vector-mediated doxycycline-inducible RNA
interference. Hum. Gene Ther. 2004, 15, 813-819
51.Palliser D., Chowdhury D., Wang Q.Y., Lee S.J., Bronson R.T., Knipe
31.Howard K.: Unlocking the money-making potential of RNAi. Nat.
D.M., Liberman J.: An siRNA-based microbicide protects mice from
lethal herpes simplex virus 2 infection. Nature 2006, 439, 89-94
32.Hua J., Mutch D.G., Herzog T.J.: Stable suppression of MDR-1 gene
52.Park W.S., Hayafune M., Miyano-Kurosaki N., Takaku H.: Specific
using siRNA expression vector to reverse drug resistance in a human
HIV-1 gene silencing by small interfering RNAs in human peripheral
uterine sarcoma cell line. Gynecol. Oncol. 2005, 98, 31-38
blood mononuclear cells. Gene Ther., 2003, 10, 2046-2050
33.Jackson A.L., Linsley P.S.: Noise amidst the silence: off-target effects
53.Putral L.N., Bywater M.J., Gu W., Saunders N.A., Gabrielli B.G.,
of siRNAs? Trends Genet. 2004, 20, 521-524
Leggatt G.R., McMillan N.A.J.: RNAi against HPV oncogenes in ce-
34.Jiang M., Arzumanov A.A., Gait M.J., Milner J.: A bi-functional siRNA
rvical cancer cells results in increased sensitivity to cisplatin. Mol.
construct induces RNA interference and also primes PCR amplifica-
tion for its own quantification. Nucleic Acids Res. 2005, 33, e151
54.Raoul C., Abbas-Terki T., Bensadoun J., Guillot S., Haase G., Szulc
35.Judge A.D., Sood V., Shaw J.R., Fang D., McClintock K., MabLachlan I.:
J., Henderson C.E., Aebischer P.: Lentiviral-mediated silencing of
Sequence-dependent stimulation of the mammalian innate immune
SOD1 through RNA interference retards disease onset and progres-
response by synthetic siRNA. Nat. Biotechnol. 2005, 23, 457-462
sion in a mouse model of ALS. Nat. Med. 2005, 11, 423-428
36.Kim. D.H., Longo M., Han Y., Lundberg P., Cantin E., Rossi J.J.: Inter-
55.Robbins M.A., Li M., Leung I., Li H., Boyer D.V., Song Y., Behlke
feron induction by siRNAs and ssRNAs synthesized by phage poly-
M.A., Rossi J.J.: Stable expression of shRNAs in human CD34+ pro-
merase. Nat. Biotechnol. 2004, 22, 321-325
genitor cells can avoid induction of interferon responses to siRNAs
in vitro. Nat. Biotechnol. 2006, 24, 566-571
37.Konstantinova P., de Vries W., Haasnoot J., ter Brake O., de Haan P.,
Berkhout B.: Inhibition of human immunodeficiency virus type 1 by RNA
56.Robinson R.: RNAi Therapeutics: How likely, how soon? PloS Biol.
interference using long-hairpin RNA. Gene Ther. 2006, 13, 1403-13
38.Kurreck J.: Antisense technologies. Improvement through novel che-
57.Ruttimann J.: RNA therapy tackles eye disease. Nature 2006, www.na-
mical modifications. J. Biochem. 2003, 270, 1628-1644
ture.com/ news/2006/060605/pf/060605-4.html
39.Lau N.C., Bartel D.P.: Cenzorzy genów. wiat Nauki, 2003, 145, 34-41
58.Saksela K.: Human viruses under attack by small inhibitory RNA.
40.Li B., Tang Q., Cheng D., Qin C., Xie F.Y., Wei Q., Xu J., Liu Y.,
Zheng B., Woodle M.C., Zhong N., Lu P.Y.: Using siRNA in prophy-
59.Sandy P., Ventura A., Jacks T.: Mammalian RNAi: a practical guide.
lactic and therapeutic regimans against SARS coronavirus in Rhesus
60.Scherr M., Eder M.: RNAi in functional genomics. Curr. Opin. Mol.
41.Li Y.C., Kong L.H., Cheng B.Z., Li K.S.: Construction of influenze
virus siRNA expression vectors and their inhibitory effects on multi-
61.Sioud M.: RNA interference below the immune radar. Nat. Biotech-
plication of influenza virus. Avian Dis. 2005, 49, 562-573
42.MacRae I.J., Zhou K., Li F., Repic A., Brooks A.N., Cande W.Z., Adams
62.Skipper M.: RNA interference. Have our dreams been shattered? Nat.
P.D., Doudna J.A.: Structural Basis for Double-Stranded RNA Pro-
cessing by Dicer. Science 2006, 311, 195-198
63.Sledz C.A., Holko M., de Veer M.J., Silverman R.H., Williams B.R.:
43.Makimura H., Mizuno T.M., Mastaitis J.W., Agami R., Mobbs C.V.:
Activation of the interferon system by short-interfering RNAs. Nat.
Reducing hypothalamic AGRP by RNA interference increases meta-
bolic rate and decreases body weight without influencing food inta-
64.Stevenson M.: Therapeutic potential of RNA interference. N. Engl. J.
65.Stipp D.: Biotechs billion dollar breakthrough. A technology called
79.Zhang W., Yang H., Kong X., Mohapatra S., Juan-Vergara H.S., Hel-
RNAi has opened the door to major new drugs. Already its revolu-
lermann G., Behera S., Singam R., Lockey R.F., Mohaptra S.S.: Inhibi-
tionizing gene research. Fortune 2003, 147, 96
tion of respiratory syncytial virus infection with intranasal siRNA na-
66.Stix G.: Kto wy³¹czy geny? wiat Nauki 2004, 159, 72-75
noparticles targeting the viral NS1 gene. Nat. Med. 2004, 11, 56-62
67.Syhan A.A., Vlassov A.V., Ilves H., Egry L., Kaspar R.L., Kazakov S.A.,
80.Zhang X.N., Xiong W., Wang J.D., Hu Y.W., Xiang L., Yuan Z.H.:
Johnston B.H.: Complete, gene-specific siRNA libraries: production
siRNA-mediated inhibition of HBV replication and expression. World
and expression in mammalian cells, RNA 2005, 11, 837-46
68.Tuschl T., Borkhardt A.: Small interfering RNAs: a revolutionary tool
81.Zhang Y., Wang Y., Gao W., Zhang R., Han X., Jia M., Guan W.:
for the analysis of gene function and gene therapy. Mol. Interv. 2002,
Transfer of siRNA against XIAP induces apoptosis and reduces tu-
mor cells growth potential in human breast cancer in vitro and in
vivo. Breast Cancer Res. Treat. 2006, 96, 267-277
69.Uprichard S.L.: The therapeutic potential of RNA interference. FEBS
82.Zhang Y., Zhang Y.F., Bryant J., Charles A., Boado R.J., Pardridge
W.M.: Intravenous RNA interference gene therapy targeting the hu-
70.Wang S., Chai Y.B., Liu F., Zhang X.Y., Jia W., Xie X., Yu W.Q., Shang
man epidermal growth factor receptor prolongs survival in intracra-
Z.C., Jin B.Q., Sun B.Z.: Effect of specific siRNA targeting against
nial brain cancer. Clin. Cancer Res. 2004, 10, 3667-3677
bcr-abl chimeric gene on chronic myelogenous leukemia cells. Zhon-
83.Zimmerman T.S., Lee A.C.H., Akinc A., Bramlage B., Bumcrot D.,
Fedoruk M.N., Harborth J., Heyes J.A., Jeffs L.B., John M., Judge A.D.,
71.Wang W., Wang C.Y., Dong J.H., Chen X., Zhang M., Zhao G.: Iden-
Lam K., McClintock K., Nechev L.V., Palmer L.R., Racie T., Rohl I.,
tification of effective siRNA against K-ras in human pancreatic can-
Seiffert S., Shanmugam S., Sood V., Soutschek J., Toudjarska I., Whe-
cer cell line MiaPaCa-2 by siRNA expression cassette. World J. Ga-
at A.J., Yaworski E., Zedalis W., Koteliansky V., Manoharan M., Vorn-
locher H.P., MacLachlan I.: RNAi-mediated gene silencing in non-
72.Wiebusch L., Truss M., Hagemeier C.: Inhibition of human cytome-
human primates. Nature 2006, 441, 111-114
galovirus replication by small interfering RNAs. J. Gen. Virol. 2004,
84.¯ukiel R., Nowak S., Wyszko E., Rolle K., Gawroñska I., Barciszew-
ska M.Z., Barciszewski J.: Suppression of human brain tumor with
73.Wiznerowicz M., Trono D.: Conditional suppression of cellular ge-
interference RNA specific for Tenascin-C. Cancer Biol. Ther. 2006,
nes: lentivirus vector-mediated drug-inducible RNA interference. J.
74.Wu C.J., Huang H.W., Liu C.Y., Hong C.F., Chan Y.L.: Inhibition of
SARS-CoV replication by siRNA. Antiviral Res. 2005, 65, 45-48
75.www.clinicaltrials.gov76.Yoon J.S., Kim S.H., Shin M.C., Hong S.K., Jung Y.T., Khang I.G.,
Shin W.S., Kim C.C., Paik S.Y.: Inhibition of herpesvirus-6B RNA
replication by short interference RNAs, J. Biochem. Mol. Biol. 2004,
77.Zentilin L., Giacca M.: In vivo transfer and expression of genes co-
ding for short interfering RNAs. Curr. Pharm. Biotechnol. 2004, 4,
78.Zhang M., Zhang X., Bai C.X., Song X.R., Chen J., Gao L., Hu J., Hong
Q.Y., West M.J., Wei M.Q.: Silencing the epidermal growth factor re-
Katedra i Klinika Neurochirurgii i Neurotraumatologii A.M.
ceptor gene with RNAi may be developed as a potential therapy for
non small lung cancer. Genet. Vaccines Ther. 2005, 30, 3-5
Activity of Meclocycline Sulfosalicylate Tested ICID 2006 Against Oral Pathogens RN JONES, TR FRITSCHE, HS SADER, PR RHOMBERG JMI Laboratories, North Liberty, Iowa, USA A M E N D E D A B S T R A C T ¥ Meclocycline SS was markedly superior to¥ Anaerobes (Table 2) were very susceptible tomeclocycline SS with 23 of 27 strains inhibited Background: Meclocycline sulfosalicylate
of the authors and THE JOURNAL and not those of Failure of Estrogen Plus Progestin Therapy for Prevention Suzanne W. Fletcher, MD, MSc The WHI is the first randomized primary prevention trialof postmenopausal hormones, and the part of the study thatcompared estrogen/progestin with placebo was terminatedearly. The data and safety monitoring board (DSMB) rec-PPROXIMATELY 38% OF POSTMENOPAUSAL
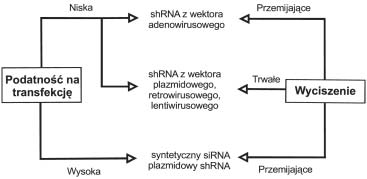 Tabela 5. Linie komórkowe wykorzystywane w badaniach
noci oraz zmian kariotypu i fizjologii (15). W zale¿noci
wyciszania genów przez siRNA (68, 63).
Tabela 5. Linie komórkowe wykorzystywane w badaniach
noci oraz zmian kariotypu i fizjologii (15). W zale¿noci
wyciszania genów przez siRNA (68, 63).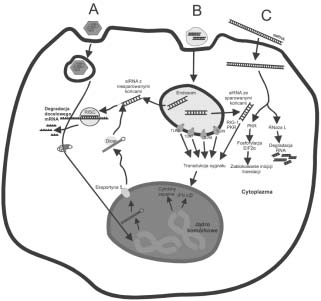 Rycina 2. Odpowied komórki na wprowadzenie ró¿nych typów dsRNA. A. Wektor wirusowy zawieraj¹cy sekwencjê shRNA inte-
gruje z genomem i ulega transkrypcji. Powsta³y shRNA jest transportowany do cytoplazmy przez eksportynê-5, hydrolizowany przez
Dicer do siRNA zawieraj¹cego niesparowane koñce i prowadzi do specyficznej degradacji docelowego mRNA. Odpowied immu-
nologiczna nie ma miejsca, gdy¿ zosta³y wykorzystane naturalne mechanizmy RNAi. B. Syntetyczny siRNA dostarczony do organi-
zmu w zawiesinie liposomalnej, dostaje siê do endosomu, gdzie aktywuje receptory TRL, co prowadzi do transdukcji sygna³u do j¹-
dra komórkowego i ekspresji genów cytokin zapalnych i interferonu. siRNA zawieraj¹cy dwa niesparowane nukleotydy przy koñcu
3 mo¿e wejæ na szlak RNAi, a siRNA ze sparowanymi koñcami aktywuje odpowied immunologiczn¹. C. Wprowadzony do ko-
mórki d³ugi dsRNA aktywuje kinazê bia³kow¹ PKR i RNazê L, co prowadzi do degradacji RNA i zablokowania inicjacji translacji,
a w konsekwencji do apoptozy. TLR ang. Toll-like receptor, PKR kinaza bia³kowa zale¿na od dsRNA, RIG-1 ang. retinoic acid-
inducible gene I, EIF2 podjednostka czynnika inicjacji translacji.
Rycina 2. Odpowied komórki na wprowadzenie ró¿nych typów dsRNA. A. Wektor wirusowy zawieraj¹cy sekwencjê shRNA inte-
gruje z genomem i ulega transkrypcji. Powsta³y shRNA jest transportowany do cytoplazmy przez eksportynê-5, hydrolizowany przez
Dicer do siRNA zawieraj¹cego niesparowane koñce i prowadzi do specyficznej degradacji docelowego mRNA. Odpowied immu-
nologiczna nie ma miejsca, gdy¿ zosta³y wykorzystane naturalne mechanizmy RNAi. B. Syntetyczny siRNA dostarczony do organi-
zmu w zawiesinie liposomalnej, dostaje siê do endosomu, gdzie aktywuje receptory TRL, co prowadzi do transdukcji sygna³u do j¹-
dra komórkowego i ekspresji genów cytokin zapalnych i interferonu. siRNA zawieraj¹cy dwa niesparowane nukleotydy przy koñcu
3 mo¿e wejæ na szlak RNAi, a siRNA ze sparowanymi koñcami aktywuje odpowied immunologiczn¹. C. Wprowadzony do ko-
mórki d³ugi dsRNA aktywuje kinazê bia³kow¹ PKR i RNazê L, co prowadzi do degradacji RNA i zablokowania inicjacji translacji,
a w konsekwencji do apoptozy. TLR ang. Toll-like receptor, PKR kinaza bia³kowa zale¿na od dsRNA, RIG-1 ang. retinoic acid-
inducible gene I, EIF2 podjednostka czynnika inicjacji translacji.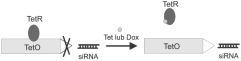 nie wektorów wirusowych daje mo¿liwoæ uzyskania sta-
bilnego wyciszenia docelowego genu, w zwi¹zku z mo¿-
liwoci¹ integracji genomu wirusa z genomem gospoda-
rza i d³ugotrwa³ej transkrypcji wyciszaj¹cego RNA. Wek-
tory wirusowe ró¿ni¹ siê miêdzy sob¹ typem preferowa-
nych komórek docelowych, ³atwoci¹ zastosowania in
Rycina 3. Zale¿noæ syntezy siRNA od tetracykliny (Tet) lub do-
vitro i in vivo, zdolnoci¹ do integracji z genomem go-
ksycykliny (Dox). TetR represor tetracyklinowy; TetO opera-
spodarza lub jej brakiem, poziomem immunogennoci
oraz maksymalnym mo¿liwym rozmiarem insertu.
nie wektorów wirusowych daje mo¿liwoæ uzyskania sta-
bilnego wyciszenia docelowego genu, w zwi¹zku z mo¿-
liwoci¹ integracji genomu wirusa z genomem gospoda-
rza i d³ugotrwa³ej transkrypcji wyciszaj¹cego RNA. Wek-
tory wirusowe ró¿ni¹ siê miêdzy sob¹ typem preferowa-
nych komórek docelowych, ³atwoci¹ zastosowania in
Rycina 3. Zale¿noæ syntezy siRNA od tetracykliny (Tet) lub do-
vitro i in vivo, zdolnoci¹ do integracji z genomem go-
ksycykliny (Dox). TetR represor tetracyklinowy; TetO opera-
spodarza lub jej brakiem, poziomem immunogennoci
oraz maksymalnym mo¿liwym rozmiarem insertu.