ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Sept. 2002, p. 2996–3000
0066-4804/02/$04.00ϩ0 DOI: 10.1128/AAC.46.9.2996–3000.2002
Copyright 2002, American Society for Microbiology. All Rights Reserved.
16S rRNA Mutation-Mediated Tetracycline Resistance in
Monique M. Gerrits,1 Marcel R. de Zoete,1 Niek L. A. Arents,2
Ernst J. Kuipers,1 and Johannes G. Kusters1*
Department of Gastroenterology and Hepatology, Erasmus MC—University Medical Center Rotterdam,Rotterdam,1 and Medical Microbiology, Regional Public Health LaboratoryGroningen/Drenthe, Groningen,2 The Netherlands
Received 12 February 2002/Returned for modification 18 April 2002/Accepted 29 May 2002
Most Helicobacter pylori strains are susceptible to tetracycline, an antibiotic commonly used for the eradi- cation of H. pylori. However, an increase in incidence of tetracycline resistance in H. pylori has recently been reported. Here the mechanism of tetracycline resistance of the first Dutch tetracycline-resistant (Tetr) H. pylori isolate (strain 181) is investigated. Twelve genes were selected from the genome sequences of H. pylori strains 26695 and J99 as potential candidate genes, based on their homology with tetracycline resistance genes in other bacteria. With the exception of the two 16S rRNA genes, none of the other putative tetracycline resistance genes was able to transfer tetracycline resistance. Genetic transformation of the Tets strain 26695 with smaller over- lapping PCR fragments of the 16S rRNA genes of strain 181, revealed that a 361-bp fragment that spanned nucleotides 711 to 1071 was sufficient to transfer resistance. Sequence analysis of the 16S rRNA genes of the Tetr strain 181, the Tets strain 26695, and four Tetr 26695 transformants showed that a single triple-base-pair substitution, AGA926-9283TTC, was present within this 361-bp fragment. This triple-base-pair substitution, present in both copies of the 16S rRNA gene of all our Tetr H. pylori transformants, resulted in an increased MIC of tetracycline that was identical to that for the Tetr strain 181. Helicobacter pylori is a spiral-shaped, gram-negative bacte-
in these genes. The second common mechanism of resistance is
rium that causes chronic infections in the gastric mucosa (6).
mediated through ribosomal protection proteins. These cyto-
This infection will persist for life, unless treated with antibiot-
plasmic proteins confer tetracycline resistance either by a re-
ics. Cure of H. pylori infection results in ulcer healing and may
duction of the affinity of ribosomes for tetracycline or by re-
reduce the risk of gastric cancer and gastric lymphoma (22, 28).
leasing the bound antibiotic from the ribosome. The ribosomal
The highest cure rates have been obtained with antimicrobial
protection proteins, such as TetM, TetO, and TetS, show ho-
treatments that include two or more antimicrobial drugs, a
mology with the elongation factors EF-G and EF-Tu (Table 1)
bismuth component, and/or a proton pump inhibitor (14, 25).
(4). Beside these two most common tetracycline resistance
For the treatment of H. pylori infections, tetracycline-based
mechanisms, two other mechanisms have been described. One
triple or quadruple therapies are often used as a second-line
is based on enzymatic inactivation of tetracycline by the prod-
treatment (7, 9, 17). Until the end of the last century only a few
uct of TetX in the presence of oxygen and NADPH, and the
reports were published on spontaneous tetracycline resistance
other originates from mutations in the 16S rRNA genes that
(18; P. D. Midolo, M. G. Korman, J. D. Turnidge, and J. R.
affect the binding site of tetracycline (4, 21, 24).
Lambert, Letter, Lancet 347:1194-1195, 1996), and it was gen-
We recently isolated a tetracycline-resistant (Tetr) H. pylori
erally accepted that tetracycline resistance (MIC Ն 4 g/ml) in
isolate (strain 181), from a 72-year-old male dyspeptic patient. H. pylori is very rare (5, 12). However, in the last 2 years an
Here we describe the molecular mechanism of tetracycline-
increase in the incidence of tetracycline resistance in H. pylori
resistance in this strain. To achieve this, 12 genes were selected
has been reported (2, 11, 13, 20, 30).
from the published H. pylori genomes (1, 23) as potential
Tetracycline inhibits the protein synthesis by binding to
candidates, based on their homology with tetracycline resis-
the 30S ribosomal subunit (3, 19). In most bacteria resis-
tance genes in other bacteria (Table 1). These putative tetra-
tance to tetracycline is due to an energy-dependent efflux of
cycline resistance genes were amplified from the genome of the
tetracycline-cation complexes across the cell membrane by
Tetr strain 181 and used for genetic transformation of the
membrane-associated efflux proteins. Export of tetracycline
tetracycline-sensitive (Tets) strain 26695 in order to identify
complexes out of the cell reduces the intracellular drug con-
the changes responsible for tetracycline resistance.
centration and protects the ribosomes from tetracycline (4).
Overexpression of the efflux genes confers tetracycline resis-
tance, while the sensitivity to tetracycline increases by deletions
MATERIALS AND METHODS Bacterial strains and growth conditions. H. pylori strains used in this study
were the Tetr strain 181 and the Tets H. pylori strains 26695 (23), J99 (1), SS1
* Corresponding author. Mailing address: Department of Gastroen-
(15), and ATCC 43504 (American Type Culture Collection). Bacteria were
terology and Hepatology, Rm. L457, Erasmus MC - University Med-
routinely grown on Columbia agar plates (Becton Dickinson, Cockeysville, Md.)
ical Center Rotterdam, Dr. Molewaterplein 40, 3015 GD Rotterdam,
supplemented with 7% lysed horse blood (BioTrading, Mijdrecht, The Nether-
The Netherlands. Phone: 31-10-4632982. Fax: 31-10-4632793. E-mail:
lands) and H. pylori Dent selective supplement (Oxoid, Basingstoke, United
Kingdom), referred to as Dent plates. Bacteria were inoculated on these plates
TETRACYCLINE RESISTANCE IN HELICOBACTER PYLORI
and incubated for 48 to 72 h at 37°C in a microaerobic atmosphere of 5% O
10% CO2, and 85% N2. Bacterial stocks were prepared by suspending bacteria,
harvested from culture plates with a sterile cotton swab, in brain heart infusion
with 20% glycerol and stored at Ϫ80°C. Determination of MIC. The MIC was routinely determined with the E-test
(AB Biodisk, Solna, Sweden) (8). Inocula were prepared from a fresh H. pylori
culture grown routinely for 2 days on Dent plates. Columbia agar plates con-
taining 7% lysed horse blood, but no Dent supplement, were inoculated with
approximately 2 ϫ 108 CFU in 20 l of 0.9% NaCl, the plates were dried for 3
to 4 min, and then the E-test strips were applied to the agar surface. The plates
were incubated at 37°C under microaerobic conditions, and 3 days later the MIC
was determined by the intercept of the zone of inhibition with the graded E-test
strip. By this method the susceptibility was determined for tetracycline, doxycy-
cline, minocycline, amoxicillin, clarithromycin, and metronidazole. The isolates
were considered resistant when the MICs of tetracycline, doxycycline, and mi-
nocycline were Ն4 g/ml and when those of amoxicillin, clarithromycin, and
metronidazole were Ն8, Ն2, and Ն8g/ml, respectively (5, 13). Natural transformation of H. pylori. Bacteria were transformed with ϳ1 g of
ϳ250 ng of PCR-amplified gene products from strain 181, as
described previously (27). Tetr transformants were selected on Dent plates con-
taining tetracycline (2 g/ml; Sigma Aldrich Chemie, Zwijndrecht, The Nether-
lands). As controls, bacteria were transformed with either genomic DNA of the
Tetr strain, TE (1 mM Tris-HCl, 0.1 mM EDTA [pH 8.0]), or DNA from the Tets
strains 26695, J99, SS1, and ATCC43504. Individual bacterial colonies present on
tetracycline-containing plates (2 g/ml) were selected, and their MICs of tetra-
PCR. Oligonucleotide primers (Isogen, Maarsen, The Netherlands) used for
TCAGAACACTTCAACCCTA TTGCTAGGCACTTCGCCATA CGCTAAAGCCTCTTGCAGTA
CCAAAAGAGCGCCAACAAAC TTTCCATTAGCGTTAGTGTC GCAGTCATTCGCTAATTCAA TCCCTTTTAGCCGCTAGTTC
PCR amplification were based on the genome sequences of H. pylori strains
26695 and J99 (Table 1; Fig. 1 and 2) (1, 23). PCR was performed in an
automated thermal cycler (I-Cycler; Bio-Rad), in a final volume of 50 l, using
the PCR-core system I (Promega, Madison, Wis.), with approximately 25 pg of
template genomic DNA and 25 pmol of each primer. Sequence analysis. Direct sequencing of the obtained PCR products was
performed by Baseclear Inc. (Leiden, The Netherlands). Sequence data were
analyzed with the help of Lasergene (DNAstar, Madison, Wis.), and Sci Ed
Central (Scientific & Educational Software, Durham, N.C.) software. Nucleotide sequence accession number. The 16S rRNA gene sequence of Tetr H. pylori strain 181 has been deposited into to the GenBank sequence database,
GTTTCCCGCTCCATTTTTA ATGGATGCGGCTAGCGATAA TGATTGGCAAAGGCCGTAGTT
CTTGCGTGTGGTAGTAATGC CTAAAGTTTTGCGCTAAGTG AACGGCTTAGCCTTATACAA ATGACATAGCCCACAAACCC
CCTACAGCTAAAGACTTGCC TTTGCCATAGAAGCTAAACG
Determination of the MICs of various antibiotics. MICs of
the four antibiotics commonly used in anti-H. pylori therapy, as
well as those of two antibiotics that belong to the tetracycline
family, were determined by E-test for the Tetr H. pylori strain181 and the H. pylori reference strain 26695 (Table 2). The
MIC of tetracycline for strain 181 was 8 g/ml (susceptibility
breakpoint Ն 4 g/ml), while the MIC for strain 26695 was
0.19 g/ml. The MICs of the two other tetracyclines, doxycy-
cline and minocycline, were also significantly higher for strain
181 than for strain 26695. For the three other routinely used
antibiotics (amoxicillin, clarithromycin, and metronidazole)
the MICs varied between Ͻ0.016 and 0.064 g/ml and did not
differ significantly between strains 181 and 26695. Transfer of tetracycline resistance by natural transforma- tion. Transformation of H. pylori strain 26695 (MIC, 0.19 g/
ml) with genomic DNA of strain 181 (MIC, 8
in Tetr colonies with a transformation frequency of 6 ϫ 10Ϫ5.
The MIC of tetracycline for the 10 randomly selected Tetr
transformants (obtained from three independent transforma-
tion experiments), determined by E-test was 8
which is identical to that for the Tetr H. pylori strain 181. The
Tetr transformants also displayed an increase of MIC of the
tetracycline derivatives, doxycycline and minocycline (Table 2). Transformation with PCR products of putative tetracycline
FIG. 1. Schematic representation of the 16S rRNA genes of H. pylori. Alignment of the 16S rRNA genes (rrnA and rrnB) from the Tetr strain
181, the Tets strain 26695, and four Tetr 26695 transformants is shown. Sequence analysis of the 16S rRNA genes revealed only a few base pair
substitutions (numbering according to 16S rrnA of H. pylori strain 26695) in the Tetr strain 181 that did not occur in the Tets strain 26695. For the
identification of the 16S rRNA region required for tetracycline resistance, the Tets strain 26695 was transformed with smaller overlapping PCR
fragments of the 16S rRNA gene of the Tetr strain 181 (only fragments containing mutations are shown). The transformants were selected on
tetracycline (2 g/ml)-containing Dent plates. Primers used for the amplification of the smaller overlapping PCR fragments started at the outside
of the fragment and each had a length of 20 bp. resistance genes. Based on their homology with tetracycline
not occur in the Tets strain 26695 (Fig. 1). Three Tetr 26695
resistance genes in other bacteria, 12 genes were selected from
transformants had incorporated the complete 16S rRNA gene
the published genome sequences of H. pylori strains 26695 (23)
of strain 181, while the fourth transformant contained the first
and J99 (1) (Table 1). The Tets H. pylori strain 26695 was
part of the 16S rRNA gene of strain 26695 and the second part
transformed with the PCR products of the selected tetracycline
of strain 181. The DNA crossover in this transformant oc-
resistance genes, which were amplified from genomic DNA of
curred after nucleotide 93 and before 127 (numbering accord-
the Tetr strain 181. Only transformation with the PCR product
ing to 16S rrnA of H. pylori strain 26695). For each strain or
of the 16S rRNA genes resulted in Tetr transformants, with a
transformant, only one sequence was obtained for the 16S
transformation frequency of 4 ϫ 10Ϫ5. No Tetr transformants
rRNA genes, indicating that these 16S rRNA genes were iden-
were found after transformation with one of the other selected
genes, TE, or DNA from the Tets strain 26695. Similar results
Identification of 16S rRNA mutations involved in tetracy-
were found for the Tets strains J99 (MIC, 0.5 g/ml), SS1
cline resistance. To determine which residues of the 16S rRNA
(MIC, 0.19 g/ml), and ATCC 43504 (MIC 0.125 g/ml). For
genes were responsible for tetracycline resistance in strain 181,
all strains the MIC of tetracycline for 10 randomly selected
the Tets strain 26695 was genetically transformed with smaller
Tetr transformants (obtained from three independent transfor-
overlapping PCR products of the 16S rRNA gene fragments,
mation experiments) determined by E-test was 8 g/ml, which
amplified from genomic DNA of the Tetr strain 181. Tetr
is identical to that for the Tetr donor strain 181.
transformants were only observed after transformation with a
Comparison of the 16S rRNA gene sequences of the Tetr
361-bp DNA fragment that spanned nucleotides 711 to 1071
strain 181, the Tets strain 26695, and four randomly Tetr 26695
(numbering according to 16S rrnA of H. pylori strain 26695),
transformants (obtained after transformation with genomic
with a transformation frequency of 5 ϫ 10Ϫ6. Transformation
DNA of strain 181) revealed several base pair differences in
with the other DNA fragments did not result in transfer of
the Tetr strain 181 as well as in the Tetr transformants that did
tetracycline resistance. The MIC of tetracycline for 10 ran-
FIG. 2. Both 16S rRNA genes are mutated in H. pylori tetracycline resistance. rrnA- and rrnB-specific sequences were amplified using specific
primers based on sequences which are found outside the two 16S rRNA genes. The rrnA-specific primers, F1 and F2, are located at position
1207020 and 1207242 (numbers corresponding to the H. pylori 26695 sequence [23]), respectively, and the rrnB-specific primers, F3 and F4, are
located at position 1510569 and 1510809, respectively. For amplification, primer R1 (located at position 1208293 and 1511828) was used in
combination with one of the other primers. All primers had a length of 20 bp.
TETRACYCLINE RESISTANCE IN HELICOBACTER PYLORI
TABLE 2. MICs for various H. pylori strains as
MIC (g/ml)a for H. pyloria Data shown are the averages of three independent experiments. The isolates
were considered resistant when the MICs of the tetracyclines (tetracycline, doxy-
cycline, and minocycline), amoxicillin, clarithromycin, and metronidazole were
Ն4, Ն8, Ն2, and Ն8 g/ml, respectively (5, 13).
domly selected Tetr transformants determined by E-test was 8
g/ml, which was identical to that for the Tetr donor strain 181.
The only difference found between these Tetr 26695 transfor-
mants and the Tets strain was the triple-base-pair substitution
Both copies of 16S rRNA genes are involved in tetracycline resistance. The primers that were originally used for amplifi-
cation of the 16S rRNA genes did not distinguish between the
two copies present on the H. pylori chromosome (1, 23). To
FIG. 3. Schematic representation of the primary binding site of
assess the involvement of each copy of the 16S rRNA genes in
tetracycline, based on the 16S rRNA structure of Thermus thermophi-
tetracycline resistance, specific oligonucleotide primers were
lus proposed by Wimberly et al. (29). The primary binding pocket for
developed (Fig. 2). These specific primers are based on se-
tetracycline is formed by the 16S rRNA residues 1054 to 1056 (box A)
quences which are located approximately 350 to 600 bp outside
and residues 1196 to 1200 (box B) of helix 34 and residues 964 to 967
of helix 31 (box C). The interactions between tetracycline and this
the both 16S rRNA genes, rrnA and rrnB. This allowed ampli-
pocket are formed by hydrophobic interactions, hydrogen bonds, and
fication of rrnA- and rrnB-specific sequences. rrnA- and rrnB-
salt bridges (3). The triple-base-pair substitution AGA926-9283TTC
containing PCR-fragments were obtained for the Tetr strain
(corresponding to bp 965 to 967 of E. coli 16S rRNA) is located in box
181, the eight 26695 transformants, and Tets strain 26695, and
their DNA sequences were determined. As expected, the rrnA
and rrnB sequences were identical, where as the sequences
outside the 16S rRNA genes were different. While the Tets
(corresponding to bp 965 to 967 of Escherichia coli 16S rRNA),
strain-derived fragments contained the AGA sequence in both
present in both copies of the 16S rRNA gene. Tetracycline has
genes, both for strain 181 and the eight 26695 transformants,
one primary and multiple secondary binding sites within the
30S ribosomal subunit (3, 19). In the primary binding site,
in both copies of the 16S rRNA genes.
tetracycline binds exclusively to the 3Ј-major domain of the 16S
rRNA. The primary binding pocket for tetracycline is formed
DISCUSSION
by the 16S rRNA residues 1054 to 1056 and 1196 to 1200 of
helix 34 and residues 964 to 967 of helix 31 (numbers corre-
Until recently tetracycline resistance in H. pylori was rare (5,
sponding to E. coli 16S rRNA) (3). The residues 1054 and 1196
12), but in the last 2 years, several Tetr H. pylori strains have
interact primarily with tetracycline through hydrophobic inter-
been isolated (2, 13, 30; Midolo et al., letter). These Tetr
actions, but the majority of the interaction with the drug is made
clinical isolates showed, besides tetracycline resistance, cross-
through hydrogen bonds and salt bridges between tetracycline
resistance to metronidazole (2, 13, 30; Midolo et al., letter).
and the 16S rRNA residues (Fig. 3) (3). In the Tetr H. pylori
The tetracycline resistance present in these strains was always
strain 181, the triple-base-pair substitution AGA926-9283TTC
transferred together with metronidazole resistance to a Tets
is located right in the primary binding site of tetracycline.
strain (13). In these Tetr strains it is not clear whether the
Mutations in this primary binding site are likely to affect the
tetracycline resistance is caused by a known metronidazole
affinity of the drug-ribosome interaction and thus the efficacy
resistance mechanism, a multidrug resistance mechanism, or
of tetracycline as a translational inhibitor.
an unknown tetracycline resistance mechanism (13). In our
In E. coli, the nucleotides G966 and C967 are located not only
Tetr H. pylori strain 181, no cross-resistance was found against
in the primary binding site of tetracycline but also in a func-
metronidazole, which indicated that the molecular mechanism
tional region of the ribosome, the P site (16, 26). Mutations in
of tetracycline resistance in strain 181 could be different from
this region may affect protein synthesis (10), either by a change
that of these earlier-described Tetr H. pylori strains.
in binding of tRNA to the P site itself or by blocking the
In H. pylori strain 181 resistance to tetracycline is mediated
conformational change needed for the tRNA binding to the A
by a single triple-base-pair substitution, AGA926-928 3 TTC
site. In H. pylori strain 181 and the Tetr transformants of strain
26695, the triple-base-pair substitution AGA
in Escherichia coli at methyl-modified sites: G966, C967, and G1207. Nucleic
no effect on the growth rate of the bacterium in the presence
Acids Res. 19:4259–4265.
11. Kim, J. J., R. Reddy, M. Lee, J. G. Kim, F. A. El Zaatari, M. S. Osato, D. Y.
or absence of tetracycline (data not shown). Similar observa-
Graham, and D. H. Kwon. 2001. Analysis of metronidazole, clarithromycin
tions were found with E. coli after the substitution of the
and tetracycline resistance of Helicobacter pylori isolates from Korea. J.
Antimicrob. Chemother. 47:459–461.
966 and C967 (10). This suggests that the triple-
12. Kusters, J. G., and E. J. Kuipers. 2001. Antibiotic resistance of Helicobacter
base-pair substitution AGA926-9283TTC present in H. pyloripylori. J. Appl. Microbiol. 90:134S-144S.
strain 181 does not seem to affect protein synthesis of H. pylori.
13. Kwon, D. H., J. J. Kim, M. Lee, Y. Yamaoka, M. Kato, M. S. Osato, F. A. El
During revision of this work, Trieber and Taylor (24) re-
Zaatari, and D. Y. Graham. 2000. Isolation and characterization of tetracy-
cline-resistant clinical isolates of Helicobacter pylori. Antimicrob. Agents
ported that an identical AGA3TTC substitution mediates tet-
Chemother. 44:3203–3205.
racycline resistance in an unrelated H. pylori strain (Midolo et
14. Laheij, R. J., L. G. Rossum, J. B. Jansen, H. Straatman, and A. L. Verbeek.
al., letter). None of the other mutations found in their isolates
1999. Evaluation of treatment regimens to cure Helicobacter pylori infection:
a meta-analysis. Aliment. Pharmacol. Ther. 13:857–864.
(G3323A, and the deletions G733 and G903 [numbering accord-
15. Lee, A., J. O’Rourke, M. C. De Ungria, B. Robertson, G. Daskalopoulos, and
ing to 16S rrnA of H. pylori strain 26695]) play a role in tetra-
M. F. Dixon. 1997. A standardized mouse model of Helicobacter pylori in-
fection: introducing the Sydney strain. Gastroenterology 112:1386–1397.
cycline resistance of the Tetr strain 181, since these mutations
16. Moazed, D., and H. F. Noller. 1990. Binding of tRNA to the ribosomal A-
were not present in our Tetr isolate. The differences found in
and P-sites protects two distinct sets of nucleotides in 16S rRNA. J. Mol.
the MIC of tetracycline for the triple-base-pair substitution
Biol. 211:135–145.
17. O’Morain, C., and S. Montague. 2000. Challenges to therapy in the future.
mutant between the study of Trieber and ours are only due to
Helicobacter 5:S23-S26.
the methods used for the determination of the MIC (data not
18. Piccolomini, R., G. Di Bonaventura, G. Catamo, F. Carbone, and M. Neri.
shown). The finding that in two unrelated H. pylori strains the
1997. Comparative evaluation of the E-test, agar dilution, and broth microdi-
lution for testing susceptibilities of Helicobacter pylori strains to 20 antimi-
exact same mutation is responsible for tetracycline resistance
crobial agents. J. Clin. Microbiol. 35:1842–1846.
opens possibilities for the development of molecular screening
19. Pioletti, M., F. Schlunzen, J. Harms, R. Zarivach, M. Gluhmann, H. Avila,
tests for tetracycline resistance in H. pylori.A. Bashan, H. Bartels, T. Auerbach, C. Jacobi, T. Hartsch, A. Yonath, and F. Franceschi. 2001. Crystal structures of complexes of the small ribosomal
subunit with tetracycline, edeine and IF3. EMBO J. 20:1829–1839.
20. Realdi, G., M. P. Dore, A. Piana, A. Atzei, M. Carta, L. Cugia, A. Manca, ACKNOWLEDGMENT B. M. Are, G. Massarelli, I. Mura, A. Maida, and D. Y. Graham. 1999.
We thank A. H. M. van Vliet for helpful comments and discussions.
Pretreatment antibiotic resistance in Helicobacter pylori infection: results of
three randomized controlled studies. Helicobacter 4:106–112.
21. Ross, J. I., E. A. Eady, J. H. Cove, and W. J. Cunliffe. 1998. 16S rRNA REFERENCES
mutation associated with tetracycline resistance in a gram-positive bacte-
1. Alm, R. A., L. S. Ling, D. T. Moir, B. L. King, E. D. Brown, P. C. Doig, D. R.
rium. Antimicrob. Agents Chemother. 42:1702–1705. Smith, B. Noonan, B. C. Guild, B. L. deJonge, G. Carmel, P. J. Tummino, A.
22. Sugiyama, T., N. Sakaki, H. Kozawa, R. Sato, T. Fujioka, K. Satoh, K. Caruso, M. Uria-Nickelsen, D. M. Mills, C. Ives, R. Gibson, D. Merberg, Sugano, H. Sekine, A. Takagi, Y. Ajioka, and T. Takizawa. 2002. Sensitivity S. D. Mills, Q. Jiang, D. E. Taylor, G. F. Vovis, and T. J. Trust. 1999.
of biopsy site in evaluating regression of gastric atrophy after Helicobacter
Genomic-sequence comparison of two unrelated isolates of the human gas-
pylori eradication treatment. Aliment. Pharmacol. Ther. 16(Suppl. 2):187–
tric pathogen Helicobacter pylori.Nature 397:176–180.
2. Boyanova, L., I. Stancheva, Z. Spassova, N. Katzarov, I. Mitov, and R.
23. Tomb, J. F., O. White, A. R. Kerlavage, R. A. Clayton, G. G. Sutton, R. D. Koumanova. 2000. Primary and combined resistance to four antimicrobial Fleischmann, K. A. Ketchum, H. P. Klenk, S. Gill, B. A. Dougherty, K.
agents in Helicobacter pylori in Sofia, Bulgaria. J. Med. Microbiol. 49:415– Nelson, J. Quackenbush, L. Zhou, E. F. Kirkness, S. Peterson, B. Loftus, D. Richardson, R. Dodson, H. G. Khalak, A. Glodek, K. McKenney, L. M.
3. Brodersen, D. E., W. M. Clemons, Jr., A. P. Carter, R. J. Morgan-Warren, Fitzegerald, N. Lee, M. D. Adams, and J. C. Venter. 1997. The complete B. T. Wimberly, and V. Ramakrishnan. 2000. The structural basis for the
genome sequence of the gastric pathogen Helicobacter pylori. Nature 388:
action of the antibiotics tetracycline, pactamycin, and hygromycin B on the
30S ribosomal subunit. Cell 103:1143–1154.
24. Trieber, C. A., and D. E. Taylor. 2002. Mutations in the 16S rRNA genes of
4. Chopra, I., and M. Roberts. 2001. Tetracycline antibiotics: mode of action, Helicobacter pylori mediate resistance to tetracycline. J. Bacteriol. 184:2131–
applications, molecular biology, and epidemiology of bacterial resistance.
Microbiol. Mol. Biol. Rev. 65:232–260.
25. van der Hulst, R. W., J. J. Keller, E. A. Rauws, and G. N. Tytgat. 1996.
5. Debets-Ossenkopp, Y. J., A. J. Herscheid, R. G. Pot, E. J. Kuipers, J. G.
Treatment of Helicobacter pylori infection: a review of the world literature. Kusters, and C. M. Vandenbroucke-Grauls. 1999. Prevalence of Helicobacter
Helicobacter 1:6–19. pylori resistance to metronidazole, clarithromycin, amoxycillin, tetracycline
26. von Ahsen, U., and H. F. Noller. 1995. Identification of bases in 16S rRNA
and trovafloxacin in The Netherlands. J. Antimicrob. Chemother. 43:511–
essential for tRNA binding at the 30S ribosomal P-site. Science 267:234–237.
27. Wang, Y., K. P. Roos, and D. E. Taylor. 1993. Transformation of Helicobacter
6. Dunn, B. E., H. Cohen, and M. J. Blaser. 1997. Helicobacter pylori. Clin. pylori by chromosomal metronidazole resistance and by a plasmid with a
Microbiol. Rev. 10:720–741.
selectable chloramphenicol resistance marker. J. Gen. Microbiol. 139:2485–
7. Gisbert, J. P., and J. M. Pajares. 2001. Helicobacter pylori therapy: first-line
options and rescue regimen. Dig. Dis. 19:134–143.
28. Wilhelmsen, I., and A. Berstad. 1994. Quality of life and relapse of duodenal
8. Glupczynski, Y., M. Labbe, W. Hansen, F. Crokaert, and E. Yourassowsky.
ulcer before and after eradication of Helicobacter pylori. Scand. J. Gastro-
1991. Evaluation of the E-test for quantitative antimicrobial susceptibility
enterol. 29:874–879.
testing of Helicobacter pylori. J. Clin. Microbiol. 29:2072–2075.
29. Wimberly, B. T., D. E. Brodersen, W. M. Clemons, Jr., R. J. Morgan-Warren,
9. Graham, D. Y., M. S. Osato, J. Hoffman, A. R. Opekun, S. Y. Anderson, D. H. A. P. Carter, C. Vonrhein, T. Hartsch, and V. Ramakrishnan. 2000. Struc- Kwon, and H. M. El Zimaity. 2000. Metronidazole containing quadruple
ture of the 30S ribosomal subunit. Nature 407:327–339.
therapy for infection with metronidazole resistant Helicobacter pylori: a pro-
30. Wu, H., X. D. Shi, H. T. Wang, and J. X. Liu. 2000. Resistance of Helicobacter
spective study. Aliment. Pharmacol. Ther. 14:745–750. pylori to metronidazole, tetracycline and amoxycillin. J. Antimicrob. Chemo-
10. Jemiolo, D. K., J. S. Taurence, and S. Giese. 1991. Mutations in 16S rRNA
ther. 46:121–123.
Westwood Evaluation & Treatment Center, 11340 Olympic Boulevard, Suite 303, Los Angeles, California 90064, 310-444-3154, [email protected] “in order to diagnose a Posttraumatic Stress Disorder correctly the doctor must show that the individual meets the DSM-IV-TR criteria” One of the most frequently encountered diagnoses in personal injury and workers’ compensation litigation
Colorado Mesa University Outdoor Program Brazil Gear List Use this to do a last minute Check! MARK YOUR INITIALS ON ALL BELONGINGS. This really helps when doing laundry. EQUIPMENT ___ Internal frame Backpack big enough to hold all your gear. ___ Duffel bag for leaving extra gear in hotels and to cover you main pack. ___ Smaller, lightweight day pack or fanny pack. (may be part of yo
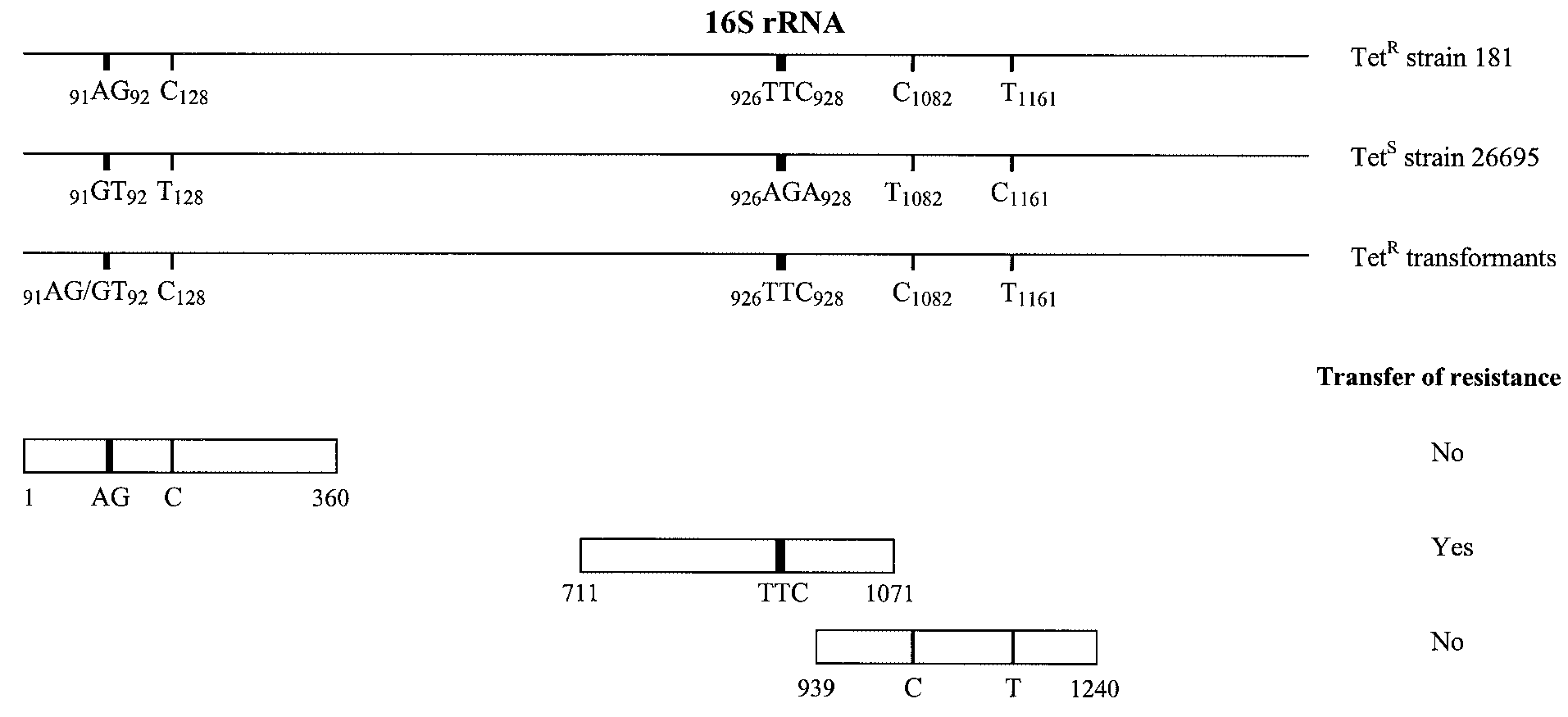
 FIG. 1. Schematic representation of the 16S rRNA genes of H. pylori. Alignment of the 16S rRNA genes (rrnA and rrnB) from the Tetr strain
181, the Tets strain 26695, and four Tetr 26695 transformants is shown. Sequence analysis of the 16S rRNA genes revealed only a few base pair
substitutions (numbering according to 16S rrnA of H. pylori strain 26695) in the Tetr strain 181 that did not occur in the Tets strain 26695. For the
identification of the 16S rRNA region required for tetracycline resistance, the Tets strain 26695 was transformed with smaller overlapping PCR
fragments of the 16S rRNA gene of the Tetr strain 181 (only fragments containing mutations are shown). The transformants were selected on
tetracycline (2 g/ml)-containing Dent plates. Primers used for the amplification of the smaller overlapping PCR fragments started at the outside
of the fragment and each had a length of 20 bp.
FIG. 1. Schematic representation of the 16S rRNA genes of H. pylori. Alignment of the 16S rRNA genes (rrnA and rrnB) from the Tetr strain
181, the Tets strain 26695, and four Tetr 26695 transformants is shown. Sequence analysis of the 16S rRNA genes revealed only a few base pair
substitutions (numbering according to 16S rrnA of H. pylori strain 26695) in the Tetr strain 181 that did not occur in the Tets strain 26695. For the
identification of the 16S rRNA region required for tetracycline resistance, the Tets strain 26695 was transformed with smaller overlapping PCR
fragments of the 16S rRNA gene of the Tetr strain 181 (only fragments containing mutations are shown). The transformants were selected on
tetracycline (2 g/ml)-containing Dent plates. Primers used for the amplification of the smaller overlapping PCR fragments started at the outside
of the fragment and each had a length of 20 bp.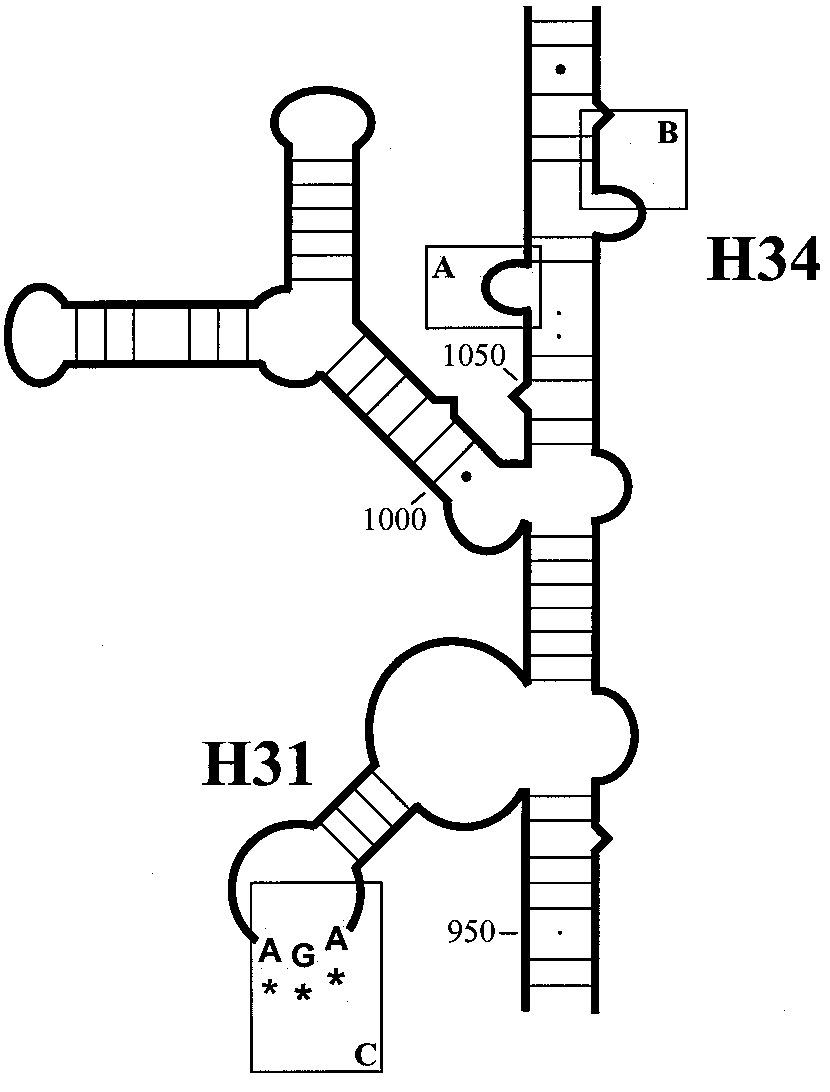 TETRACYCLINE RESISTANCE IN HELICOBACTER PYLORI
TABLE 2. MICs for various H. pylori strains as
MIC (g/ml)a for H. pylori
a Data shown are the averages of three independent experiments. The isolates
were considered resistant when the MICs of the tetracyclines (tetracycline, doxy-
cycline, and minocycline), amoxicillin, clarithromycin, and metronidazole were
Ն4, Ն8, Ն2, and Ն8 g/ml, respectively (5, 13).
TETRACYCLINE RESISTANCE IN HELICOBACTER PYLORI
TABLE 2. MICs for various H. pylori strains as
MIC (g/ml)a for H. pylori
a Data shown are the averages of three independent experiments. The isolates
were considered resistant when the MICs of the tetracyclines (tetracycline, doxy-
cycline, and minocycline), amoxicillin, clarithromycin, and metronidazole were
Ն4, Ն8, Ն2, and Ն8 g/ml, respectively (5, 13).