JOURNAL OF BACTERIOLOGY, Sept. 1997, p. 5827–5834
0021-9193/97/$04.00ϩ0Copyright 1997, American Society for Microbiology
A Tn10 Derivative (T-POP) for Isolation of Insertions with
Conditional (Tetracycline-Dependent) Phenotypes
Department of Biology, University of Utah, Salt Lake City, Utah 84112
Received 29 January 1997/Accepted 14 July 1997
A new Tn10-based transposon has been constructed and used to isolate insertion mutations with tetracy- cline-conditional phenotypes. Classes of mutants include conditional lethal mutations, conditional auxotrophs, and conditional mutants of the eut (ethanolamine utilization) operon. The described mutations were made with a new derivative of Tn10dTet that we have called Tn10d(T-POP). Others have noted that transposon Tn10dTet directs weak tetracycline-inducible transcripts out of both ends of the element into adjacent sequences. We have increased this level of outward transcription from Tn10dTet by selecting deletion mutations within the element that presumably remove transcription-termination signals. Insertion of the Tn10d(T-POP) element within an operon disrupts the target gene and makes expression of distal genes dependent on induction of outward transcription by tetracycline. Insertion mutations made with Tn10d(T-POP) can cause tetracycline- correctable conditional phenotypes based on expression of distal genes.
Transposable elements have been widely used in bacterial
In this paper, we describe Tn10dTet derivatives (called T-
genetics. The most commonly used elements are derived from
POP) that provide higher regulated expression of adjacent
transposon Tn10 or from bacteriophage Mu (for a review, see
genes due to deletion mutations that allow more tetracycline-
reference 19). Insertion of these elements within an operon has
induced transcription to proceed out of the transposon. Isola-
an extremely strong polar effect on the expression of genes
tion of Tn10d(T-POP) insertions between any gene and its
downstream of the insertion site. This absolute polarity limits
promoter renders expression of that gene dependent on tetra-
the usefulness of transposon insertion mutations in comple-
cycline. These new Tn10d(T-POP) elements should prove use-
mentation tests because multiple genes can be inactivated.
ful in genetic analysis since they avoid the problems of polarity
Such polarity effects likewise prevent recovery of insertions in
and allow isolation of single-gene null insertion mutations in
operons that include essential downstream genes. These limi-
tations would be removed by a transposon that provided forthe expression of distal genes. MATERIALS AND METHODS
The Tn10dTet element is a derivative of transposon Tn10
made by introducing two deletions which remove the inside
Bacterial strains. All strains are derivatives of Salmonella typhimurium LT2
(Table 1). The Tn10dTet element is a transposition-defective derivative of trans-
ends of both IS10 elements and eliminate both transposase
poson Tn10 from which the transposase gene and internal ends of IS10 have
genes (31). The deletions leave intact the outside IS10 end
been deleted (31). The MudA element is a transposition-defective derivative of
sequences (transposase substrates) and the central region
phage Mu that provides ampicillin resistance and carries a lacZYA operon for
which encodes resistance to tetracycline. This central region
creating transcriptional fusions (6, 18). The MudJ element is a transposition-defective derivative of phage Mu described by Castilho et al. (7) encoding
includes two genes, tetA and tetR, which are transcribed from
resistance to kanamycin. Plasmids encoding Tn10 transposase were kindly pro-
divergent tetracycline-inducible promoters. The tetA gene en-
codes a tetracycline efflux protein (for a review on Tn10-based
Media. Complex medium was nutrient broth (NB; 0.8%; Difco Laboratories)
tetracycline resistance, see reference 16). The tetR gene en-
supplemented with NaCl (0.5%). Minimal medium was Vogel and Bonner Emedium (12) with added glucose (0.2%) as a carbon and energy source. When
codes a regulatory protein which prevents transcription from
analyzing the metabolism of carbon sources other than glucose, minimal E
the tetA and tetR promoters in the absence of tetracycline.
medium lacking citrate (NCE) (3) supplemented with the appropriate carbon
Tetracycline interacts with the TetR protein to relieve repres-
source (0.2%) was used. When required, nutritional supplements were added to
sion and induce transcription of both the tetR gene and the tetA
E and NCE media at previously recommended final concentrations (12). Thechromogenic -galactosidase substrate 5-bromo-4-chloro-3-indolyl--D-galacto-
pyranoside (X-Gal) was added to solid minimal lactose medium (final concen-
Studies of the rnc operon using the Tn10dTet element indi-
tration, 20 g/ml) to help visualize Lacϩ colonies. Antibiotics were added at the
cated that transcripts initiated at the tet promoters extend
following concentrations: tetracycline, 20 g/ml (in complex medium) or 10
beyond the boundaries of this element and provide low-level
g/ml (in minimal medium); kanamycin, 50 g/ml; ampicillin, 30 g/ml forsingle-copy resistance determinants (MudA elements) or 100 g/ml when main-
regulated transcription of adjacent genes (30). This conclusion
taining multicopy plasmids with resistance determinants. When selecting LacTD
was based on the fact that an insertion of Tn10dTet within the
mutants (tetracycline-dependent Lacϩ; see below), tetracycline was used at a
rnc operon allows tetracycline-dependent expression of the
final concentration of 2 g/ml. Solid media were prepared by the addition of agar
distal, essential era gene. However, the level of outward tran-
(1.5%; Difco) to NB or minimal medium. Genetic techniques. Transductional crosses were mediated by the high-fre-
scription from Tn10dTet, as measured by lacZ operon fusion
quency generalized transducing phage mutant P22 HT105/1 int-201 (27). Trans-
ductants were single colony purified and made phage-free by streaking on non-selective green indicator plates (8). Cross-streaking to check phage sensitivitywas done with a P22 clear-plaque mutant, H5. Selection of mutants that relieve termination of transcription from Tn10dTet.
* Corresponding author. Mailing address: Department of Biology,
A Mud-lac insertion was placed downstream of a Tn10dTet insertion within
University of Utah, Salt Lake City, UT 84112. Phone: (801) 581-3412.
either the eut or the his operon. The orientation of the Tn10dTet element was
Fax: (801) 585-6207. E-mail: [email protected].
determined by PCR as described below. Selection for mutants that expressed the
† Present address: Department of Biology, University of California,
lac genes of the Mud-lac reporter was carried out by plating 0.1 ml (approxi-
mately 108 cells) from independent, saturated NB cultures onto minimal NCE–
ductants are due to transposition of the Tn10dTet element from the transducedparticle. To test the ability of the MudJ element to transpose, lysates from the
Lacϩ mutant strains were used in crosses with recipient strain TT8353, whichexpresses the MuA and MuB genes from plasmid pLP103-6-3 (obtained from P.
Van de Putte). Kanamycin-resistant transductants with a Hisϩ phenotype indi-
cated Mud transposition to a new chromosomal site.
Wild-type LT2/pLP103-6-3 (Ampr MuAϩ MuBϩ)
Reconstruction experiments were done to determine if the LacTD phenotype
resulted from modification of the Tn10dTet element or from unlinked mutations
Wild-type LT2/pNK972 (Ampr tnpAϩ)
that relieved transcription termination. By use of the mutant strains as donors,
transductional crosses were done to recombine the Tn10dTet insertions into a
recA1/pZT380 (Ampr lacI ptac-tnpA)
clean MudJ background and to determine the linkage of the LacTD phenotype to
Wild-type LT2/pNK2881 (Ampr tnpA*)
the Tetr phenotype of the Tn10dTet element. Final evidence that the mutations
eutA208::Tn10dTet (R) eutB8::MudA
were confined to the Tn10dTet element was obtained by transposing the
eutA208::Tn10dTet[del20] (R) eutB8::MudA
Tn10dTet element out of its original insertion site and then transposing it back
eutA208::Tn10dTet[del21] (R) eutB8::MudA
into the original site in the his operon, a hot spot for Tn10 insertion (15). The
eutA208::Tn10dTet[del22] (R) eutB8::MudA
first transposition event was obtained by a transductional cross into the recArecipient strain TT14545 (as described above). The resultant strain (carrying an
eutA208::Tn10dTet[del23] (R) eutB8::MudA
insertion of the element at an unknown site) was then used to create a random
hisG10175::Tn10dTet (A) hisC9955::MudJ
pool of Tn10dTet derivative insertion mutants in a hisϩ background by a second
hisG10175::Tn10dTet[del24] (A) hisC9955::MudJ
cross into strain TT14545. This pool (containing at least 20,000 independent
hisG10175::Tn10dTet[del25] (A) hisC9955::MudJ
transposition events) was screened for HisϪ insertions as outlined below. hisG10175::Tn10dTet[del26] (A) hisC9955::MudJ
The Tetr transductants were pooled, and the free phage titer was reduced by
hisG10175::Tn10dTet[del27] (A) hisC9955::MudJ
washing the cells twice with E salts. A P22 lysate was prepared on this pool and
hisG10175::Tn10dTet[del28] (A) hisC9955::MudJ
used to transduce wild-type LT2. The resultant Tetr transductants were replica-
hisG10176::Tn10dTet[del25] (R) hisC9955::MudJ eut-240
printed to identify HisϪ auxotrophs. Because the hisG gene includes a major hot
hisG10175::Tn10dTet[del20,del25] (R)
spot for Tn10 insertion (15), roughly half of the HisϪ insertions are at this single
site within the hisG gene. This made it possible to return a mutant element (in
hisC9955::MudJ eut-240(eut-cysA deletion)
either orientation) to the same site at which it originated. recA1 zzz-3831::Tn10Tet[del20,del25]
Determining the orientation and position of Tn10dTet insertions. To deter- proAB47/FЈ128 (pro lac) zzf-3832::Tn10Tet[del20]
mine which tet promoter was directed towards the Mud-lac reporter fusion,
proAB47/FЈ128 (pro lac) zzf-3833::Tn10Tet[del25]
Tn10dTet insertions were oriented by the PCR. These reactions were run by use
proAB47/FЈ128 (pro lac) zzf-3834::Tn10Tet[del20,del25]
of an internal Tn10dTet primer specific for one side of the element and a second
primer specific for a neighboring hisG or eutB sequence. This allowed determi-
hisG10180::Tn10dTet[del20,del25] (A) hisC9955::MudJ
nation of both the orientation of the insertions and their physical location within
hisG10181::Tn10dTet[del20,del25] (R) hisC9955::MudJ
the his or eut operons. Molecular characterization of Tn10dTet derivatives. A PCR was performed hisG10182::Tn10dTet[del20,del25] (R) hisC9955::MudJ
with primers internal to the Tn10dTet element to investigate changes in the
hisD10183::Tn10dTet[del20,del25] (A) hisC9955::MudJ
regions of Tn10dTet distal to the tetR and tetA coding regions. The PCR products
hisD10184::Tn10dTet[del20,del25] (R) hisC9955::MudJ
obtained from these reactions were purified with the Wizard PCR purification kit
hisD10185::Tn10dTet[del20,del25] (R) hisC9955::MudJ
(Promega) and sequenced with the Mn2ϩ Reagent Kit for DNA Sequencing
hisD10186::Tn10dTet[del20,del25] (R) hisC9955::MudJ
(Sequenase version 2.0, kit no. 70130; U.S. Biochemicals). hisC10187::Tn10dTet[del20,del25] (R) hisC9955::MudJ
Placement of Tn10dTet derivatives on F episomes. To construct large pools of recA1 zzz-3839::Tn10dTet[del20,del26]
random insertion events, the Tn10dTet derivatives were moved onto FЈ plasmids
proAB47/FЈ128 (pro lac) zzf-3840::Tn10dTet[del26]
by transposition. The FЈ episome carries the pro and lac genes and was main-tained by selection for prototrophy in strains carrying chromosomal mutations in
proAB47/FЈ128 (pro lac) zzf-3841::Tn10dTet[del20,del26]
the proA (TR5656) or the proAB (TR1810) genes. Chromosomal Tn10dTet
elements were transduced into a strain (TR1810) carrying the FЈ(pro lac) plas-
hlpA1::Tn10dTet[del20,del25]
mid. We then introduced into this strain plasmid pZT380, which carries an IPTG
purB2372::Tn10d(T-POP) PurTD
(isopropyl--D-thiogalactopyranoside)-inducible Tn10 transposase. The addition
pyrE2822::Tn10d(T-POP) PyrTD
of IPTG (1.0 mM) to an exponentially growing culture of the resultant strain
serA1477::Tn10d(T-POP) SerTD
induces transposition of the chromosomal Tn10dTet insertion. Following the
purB2371::Tn10d(T-POP) PurTD
addition of IPTG, the culture was grown to saturation and the FЈ element was
serA1478::Tn10d(T-POP) SerTD
moved to a new strain (TR5656) by conjugation, with selection for Proϩ andcounterselection of the donor with streptomycin. Exconjugants (Proϩ Strr) were
a Tn10dTet insertion orientation relative to downstream mud fusion: (R), tetR
screened for tetracycline resistance to identify those that carry a Tn10dTet
gene transcribed toward Mud-lac fusion; (A), tetA gene transcribed toward
insertion on the transferred FЈ episome. Strains carrying this FЈ plasmid with the
Mud-lac fusion. TD (superscript), tetracycline dependent.
Tn10dTet element were used as donors in subsequent transposition experiments;because the FЈ plasmid is of Escherichia coli origin, it does not recombine withthe Salmonella chromosome and the Tn10dTet element can be transductionallyinherited only by transposition.
lactose–X-Gal medium containing tetracycline (2 g/ml). Tetracycline was in-
Determination of Tn10d(T-POP) insertion sites. Sequences adjacent to the
cluded to induce the tet promoters, allowing isolation of mutants in which
Tn10d(T-POP) insertion in the hlpA gene were amplified by single-primer PCR
transcription of the lac genes was dependent on tetracycline. Mutant Lacϩ
done under low-stringency conditions (45 cycles of 1 min at 94°C, 1 min at 45°C,
colonies arose on the plates after 2 days at 37°C, with additional mutants ap-
and 45 min at 72°C; Idaho Technologies Thermocycler). Internal, outward-
pearing upon additional incubation for up to 4 days. These Lacϩ colonies were
directed primers specific to each side of the Tn10d(T-POP) element were used
tested to determine whether their Lacϩ phenotype depended on the presence of
individually for this initial PCR. PCR products having one endpoint within the
Tn10d(T-POP) element were identified by comparison with the bands produced
Genetic characterization of LacTD mutants. Transductional linkage between
with template DNA from an isogenic strain lacking the Tn10d(T-POP) element.
the Tetr phenotype of the Tn10dTet element and the drug resistance of the
The unique PCR products were excised and purified with the QIAquick gel
downstream Mud element (Kanr for MudJ; Ampr for MudA) was tested. Link-
extraction kit. The sequences were reamplified by high-stringency PCR with a
ages substantially greater than those observed for the parent strain indicated that
nested primer within Tn10d(T-POP) used in 100-fold excess with the original
the Lacϩ mutant strain contained a deletion of the intervening region. For
primer used for single-primer PCR. The final, somewhat-smaller PCR product
mutant strains obtained from the hisG::Tn10dTet insertion, a loss of hisD gene
was purified and sequenced as described above. This was done for both sides of
expression (required for growth on minimal histidinol medium) also indicated
Isolation of Tn10d(T-POP) insertions in the eut operon. New insertions of the
Any mutation (deletion or point mutation) that altered or removed the trans-
Tn10d(T-POP) element in the eut operon were isolated by transducing the
posase binding sites at the ends of a Tn10dTet element or MudJ element would
Tn10d(T-POP) element (from strain TT18797) into a recipient strain expressing
be expected to eliminate the ability to transpose. To test for Tn10dTet transpo-
the Tn10 transposase from plasmid pNK2881 (19). The recipient also carried a
sition, P22 lysates prepared on the Lacϩ mutant strains were used as donors in
Mud-lac fusion element in the last gene of the eut operon (eutR); the Mud-lac
a cross with a RecAϪ recipient strain (TT14545) that expresses the Tn10 trans-
fusion is not expressed since eut operon transcription depends on the EutR
posase from plasmid pNK972. The recipient recA mutation prevents inheritance
protein (26). Insertions of the Tn10d(T-POP) element were isolated as Lacϩ
of the donor Tn10dTet insertion by homologous recombination; all Tetr trans-
transductants on minimal NCE-lactose-tetracycline medium and shown to be
FIG. 1. Selection systems for modification of Tn10dTet. These abbreviated maps show the positions of Tn10dTet and MudJ insertions within the ethanolamine (eut)
(A) and histidine (his) (B) operons of the parent strains. The eutA and eutB genes are the 11th and 12th genes in the ethanolamine operon. The hisG, -D, and -C genesare the first three genes in the histidine operon. The hisG::Tn10dTet insertion is at a hot spot for Tn10 insertion.
LacϪ on medium lacking tetracycline. These candidate insertions were then
lactose-tetracycline medium. In the presence of tetracycline,
transduced into eutRϩ backgrounds to test their Eut phenotype with and without
the parental strain TT18784 containing the hisG::Tn10dTet
insertion became HisDϩ, indicating that the low level of tet-racycline-induced transcription of hisD from the parental
transposon is sufficient to produce histidine from histidinol. Modification of left and right ends of Tn10dTet. Insertions
Mutants that expressed the lac genes to a much higher level
of Tn10dTet were placed upstream of Mud-lac insertions, and
were selected on minimal lactose medium with tetracycline.
the double mutant strains were used to select Lacϩ derivatives
Revertants (Lacϩ) appeared at a frequency of 10Ϫ6 to 10Ϫ7/
with increased transcription extending from the ends of the
cell plated. The majority of the revertants were shown to be
Tn10dTet element into the Mud-lac element. Modification of
Lacϩ with or without tetracycline. The LacTD strains were
the left (tetR) end of Tn10dTet was selected by use of inser-
tested for linkage between the Tn10 and MudJ elements and
tions in the eut operon (Fig. 1A). The Tn10dTet insertion was
for the ability of the Tn10 element to transpose. Eight inde-
in the eutA gene and oriented so that tetR transcription pro-
pendent LacTD mutants from the eutA::Tn10dTet insertion
ceeds in the same direction as that of the eut operon. The
and 16 from the hisG::Tn10dTet insertion were analyzed and
MudA element was located in the eutB gene immediately
13 of them behaved as expected for having mutations within
downstream. The Tn10dTet element blocks transcription of
the Tn10 element. Classification of the LacTD mutants is de-
the MudA lac operon from the main eut promoter; inducedtranscription from Tn10dTet is insufficient to provide a Lacϩphenotype. The Mud-lac fusion is not expressed unless a new
TABLE 2. Mutant classes obtained in LacTD selection
mutation provides for increased transcription.
For mutations affecting the right (tetA) end of Tn10dTet, a
similar system was used (Fig. 1B) but in the his operon; the
Tn10dTet element was located in the promoter-proximal hisG
gene and was oriented such that the tetA gene is transcribed in
the same direction as that of the his operon. The MudJ inser-
tion was located in the downstream hisC gene. Transcription
from the hisG::Tn10dTet element could be monitored by ex-
Mutations internal to Tn10dTetd
pression of the hisC::MudJ lac genes or by expression of the
intervening hisD gene. Since the HisD enzyme converts his-
tidinol to histidine, its expression allows a HisϪ strain to grow
Assay of -galactosidase activity allowed quantification
Mutations internal to Tn10dTetd
of the tetracycline-induced transcription emerging from the
Tn10dTet element. For the original Tn10dTet element, the
Total number of LacTD mutant classes from 10 independent saturated cul-
tures of the parent strain TT18778. Mutations characterized in footnotes b to d
addition of tetracycline to the growth medium results in a 4.5-
are a subset of these LacTD mutants.
to 5-fold increased transcription of the Mud lac reporter ad-
b Tn10dTet and MudJ elements were 100% linked (inseparable in transduc-
jacent to the tetR or tetA ends of (see Table 3, rows 1 and 6,
respectively); this was originally demonstrated by Takiff et al.
The Tn10dTet element was unable to transpose when provided with trans-
(30). However, the induced levels of -galactosidase activity
d LacTD mutation showed 100% linkage to the Tn10dTet element. The
are insufficient to allow the parental strains to grow on minimal
Tn10dTet element was able to transpose when provided with transposase.
FIG. 2. Position of deletion mutations within Tn10dTet. Deletions at the tetR end (left) were selected by use of the insertions in the ethanolamine operon; those
at the tetA end (right) were selected by use of the insertions in the histidine operon. Numbers indicate the positions of each feature in the DNA base sequence of theparent transposon, Tn10dTet.
scribed in Materials and Methods, and the results are summa-
(Fig. 2, del24 to del28). The published sequence (17) shows a
rized in Table 2. Six eutA::Tn10dTet mutants and five
stem-loop structure, characteristic of rho-independent termi-
hisG::Tn10dTet mutants were sequenced; the chosen strains
nators, just downstream of the tetA gene. Although this struc-
were those showing the greatest induced expression of lacZ.
ture seemed a likely candidate site for mutations that allow
Based on the inducibility of the Lacϩ phenotype and the
increased transcription out of the right end of Tn10dTet, only
tetracycline resistance, the LacTD mutations were expected to
two of the five deletions (del24 and del26) removed this puta-
lie outside of the tetR and tetA coding regions. Oligonucleo-
tides were designed to prime PCR amplification of the regions
Quantification of transcription levels from Tn10dTet deriva-
between the tetR or tetA gene and the IS10 ends. All PCR
tives. Transcription extending from the derivatives of Tn10dTet
products obtained from the mutant strains were smaller than
was determined by -galactosidase assays. Without tetracy-
those from the respective parent strains, indicating that the
cline, transcription from both tet promoters is essentially the
LacTD phenotype resulted from deletions within the Tn10dTet
element. Sequencing of the PCR products and comparison of
the sequence to the parental sequence localized the endpoints.
parent strains (Table 3, column 5), which is consistent with the
Five of the six tetR end deletions were characterized. Two of
tight repression previously described for the tet promoters (5,
the deletions appeared to be identical, leaving four different
10). The addition of tetracycline dramatically increased tran-
deletions (Fig. 2, del20 to del23). As expected, none of the
scription of downstream lacZ reporter genes to levels well over
deletions disrupts the tetR coding sequence. Two deletions
300 -galactosidase units (Table 3, column 6). For many of the
(del20 and del23) extend into IS10 material, but neither re-
strains containing deletions within the Tn10dTet elements, ex-
moves the end sequences necessary for transposition. The col-
pression was induced at least 30-fold, reaching a level 5- to
lection of deletions at the tetA side was more heterogeneous
10-fold higher than the induced level of the parent strains
TABLE 3. Expression of eut::MudA and his::MudJ lac fusions from Tn10 promoters
a Tn10dTet insertion orientation relative to downstream Mud fusions: (R), tetR gene transcribed towards Mud-lac fusion; (A), tetA gene transcribed towards Mud-lacb Transcription of downstream lac fusion (in Miller units). For eut::Tn10dTet insertions, a eutB::MudA element was used, for his::Tn10dTet insertions, a hisC::MudJ
c Inducing tetracycline was provided in growth medium at 2.0 g/ml.
FIG. 3. Construction of a transposon with two deletion mutations. The diagrammed cross was used to construct T-POP, the double deletion mutant Tn10dTet
(del25, del20). The recipient carries mutation del25 but is oriented such that the end needed for expression of the MudJ lac genes has no deletion; this strain isphenotypically LacϪ. The donor strain has mutation del20 at the end nearest the MudA-lac operon. Therefore, this strain is phenotypically LacTD. In the cross, selectionis made for LacTD recombinants; this demands inheritance of the donor mutation del20 (events allowing this are indicated by roman numerals). The donor elementscannot be inherited at the recipient eut locus because it carries a large eut deletion mutation. All recombination events must occur within the two transposons sequencesshared by donor and recipient.
(Table 3, compare rows 1 and 6 to rows 2 to 5 and rows 7 to
Tn10dTet element was moved out of the his operon and sub-
sequently onto an FЈ plasmid by transposition as described in
Construction of recombinant Tn10dTet derivatives with a
Materials and Methods. The element carrying both deletions
deletion at both ends of the element. A transductional cross
(Tn10dTet[del20,del25]) has been named Tn10d(T-POP) or
was done to form the desired double-mutant element by re-
T-POP for the fact it confers (t)etracycline resistance and
combination between two single-mutant Tn10dTet elements,
utilizes the divergent (p)romoter-(o)perator-(p)romoter sys-
one in the eutA gene and one in the hisG gene. This cross is
tem for providing expression of adjacent genes. By similar
diagrammed in Fig. 3. The donor strain (TT18779) carries a
manipulations, a second double-mutant element that carries
eutA::Tn10dTet[del20] insertion and a eutB::MudA(lac Ampr)
the same tetR end deletion (del20) in combination with a dif-
insertion; this strain is phenotypically Lacϩ in the presence of
ferent tetA end deletion (del26) has been constructed; the
tetracycline. The recipient strain (TT18792) contains an inser-
second element has been designated T-POP2.
tion of the Tn10dTet[del25] element in the hisG gene with its
Characterization of his mutants generated by the Tn10d(T-
unmodified (tetR) end oriented towards the hisC::MudJ ele-
POP) insertion. Insertions of the new Tn10d(T-POP) element
ment; this strain is phenotypically LacϪ. The transduction was
at new sites in the his operon were isolated by screening ap-
done by selecting for Lacϩ recombinants on minimal lactose-
proximately 35,000 independent insertion mutants. These HisϪ
tetracycline medium supplemented with histidine and cystine.
insertions were mapped by a set of his deletions to identify
The Lacϩ donor and the LacϪ recipient strains have different
insertions in the hisGDC region.
sequences flanking their Tn10dTet and Mud elements (either
Each of these new insertions of Tn10d(T-POP) was moved
eut or his), and recombination in the eut region is prevented by
into the hisC::MudJ background by transduction. The location
the eut-cysA deletion in the recipient. Therefore, all Lacϩ re-
and orientation of the new insertions were determined more
combinants must arise by exchange between donor and recip-
precisely by PCR. Based on the strong insertion site specificity
ient Tn10dTet and Mud transposons. The desired recombinant
of the Tn10 transposase (15), strains producing similarly sized
type with deletions at each end is expected to have both theTn10dTet and Mud elements within the his operon, separatedby an intervening eut sequence.
Lacϩ transductants were isolated and tested for the LacTD
phenotype. All recombinants were HolϪ (even with tetracy-cline) due to loss of the hisD gene, and all showed 100%linkage of the Tetr and Kanr phenotypes since nonhomology ofthe intervening eut sequence prevents recombination betweenthe two elements. The Tn10 elements of several Lacϩ trans-ductants were screened by PCR for possession of the deletionsat either side of the Tn10dTet. A recombinant (TT18793)
FIG. 4. Positions of T-POP insertions in the histidine operon. Random in-
sertions in the operon were isolated to test the behavior of T-POP at a variety of
which showed the bands predicted for each of the two dele-
sites. Positions were estimated by PCR. The behavior of these insertions is
tions was saved for later use (data not shown). This composite
TABLE 4. Transcription levels from his::Tn10d(T-POP) insertions
a few his sites were tested and that the method used for thosesites is not very sensitive, since low levels of HisD enzyme are
sufficient for growth. However, in many more tests of the eut
operon, we have seen no evidence of antisense effects on up-
stream gene expression (data not shown). Isolation and characterization of conditional mutations gen- erated by Tn10d(T-POP) insertion. To test the general useful-
ness of the Tn10d(T-POP) element, other chromosomal inser-
tions were isolated from a pool of random Tn10d(T-POP)
insertions. A lysate prepared on this pool of insertions was
used to transduce LT2 on NB-tetracycline medium. Tetracy-
cline-resistant transductants were replica-printed to minimal
E-glucose medium lacking tetracycline to identify insertion
a Tn10d(T-POP) insertions orientation relative to downstream Med fusion:
mutants that caused auxotrophy or lethality in the absence of
(A) tetA promoter directed towards MudJ insertion (R), tetR promoter directed
tetracycline. From this initial screen, 21 Tn10d(T-POP) inser-
tions were isolated. The classification of these mutants is given
Transcription of downstream lac fusion (in Miller units); a hisC::MudJ ele-
in Table 5. This same general distribution of mutant types has
c Tetracycline was provided in growth medium at 2.0 g/ml.
been observed for a larger set of Tn10d(T-POP) insertionsisolated by a genetics laboratory course. We mapped the mu-tants whose growth phenotype was corrected by tetracycline;
PCR products are likely to be insertions at the same site. A set
the mapping method was that of Benson and Goldman (2).
of Tn10d(T-POP) insertions is diagrammed in Fig. 4. These
One Tn10d(T-POP) insertion (TT19197) exhibited a tetra-
strains were chosen based on their having either a distinct in-
cycline-correctable, lethal phenotype and mapped to the 4- to
sertion site or a distinct orientation. Tetracycline-induced tran-
7-min region of the Salmonella chromosome. To identify the
scription levels from these Tn10d(T-POP) elements are pre-
exact location of this conditional lethal Tn10d(T-POP) inser-
sented in Table 4. Strains in which the lacZ gene is expressed
tion, a low-stringency single-primer PCR (as described in Ma-
from the tetR promoter showed variability in the levels of tran-
terials and Methods) was used to amplify DNA adjacent to
scription depending on the insertion site. Insertion of Tn10d
the insertion site for sequencing. A FASTA analysis of the
(T-POP) at the hisD10184 site exhibited high levels of induced
sequences flanking the Tn10d(T-POP) insertion with se-
transcription (Table 4, row 5), yet insertion at the hisG10182
quences from the E. coli genome database indicated that the
site generated virtually no tetracycline-induced expression of
Tn10d(T-POP) insertion was located in the Salmonella ho-
-galactosidase (Table 4, row 4). In contrast, transcription from
molog of the E. coli hlpA gene. Alignments of the Salmonella
the tetA promoter consistently showed a high level of induction
sequence on either side of the Tn10d(T-POP) element and the
for his operon insertions (Table 4, rows 1 and 2) as well as for
E. coli hlpA sequence showed 84 and 85% nucleotide sequence
insertions obtained in other operons (data not shown).
identity. Furthermore, the hlpA gene is located at min 4.4 of
Investigation of the effects of antisense transcripts from Tn10d
the E. coli genetic map (4). The lethal phenotype of the hlpA(T-POP) elements. With an element that produces transcripts
insertion is likely due to a polar effect on expression of essen-
extending out of both ends, the possibility exists that the mes-
tial downstream genes; this expression could be restored by the
sage directed upstream (opposite that of the target operon)
addition of tetracycline. The hlpA gene is part of an operon
reduces expression of genes upstream of the insertion site. To
that includes essential functions required for the synthesis of
investigate this, we examined the HisD phenotype following
lipid A, the hydrophobic anchor for lipopolysaccharides in the
induction of transcription from a Tn10d(T-POP) element in-
outer membrane (for reviews, see references 22 and 28).
serted in the downstream hisC gene (TT19186). Strain TT19186
Thus, the use of the T-POP element provides in vivo evidence
showed no difference in growth on minimal E medium-glu-
that the hlpA gene is dispensable but is located within an
cose-histidinol plates than on identical plates with added tetra-
operon that includes distal genes for essential proteins.
cycline (2.0 g/ml). This result and those obtained from Tn10d
Identification of phenotypically silent genes in the eut oper-
(T-POP) insertions in the eut operon (see below) show that
on. Genetic analysis of the ethanolamine utilization operon
antisense messages from the Tn10d(T-POP) element do not
(eut) of Salmonella indicated six genes whose mutation caused
cause major interference with the expression of genes up-
an aerobic EutϪ phenotype (25). Subsequent sequence deter-
stream of the insertion site. It should be kept in mind that only
mination (14, 21, 29) has revealed 16 genes in the operon,
TABLE 5. Characterization of Tn10d(T-POP) insertions
2 histidine auxotrophs, 2 tryptophan auxotrophs, 2 leucine auxotrophs, 2 proline auxotrophs,
1 methionine auxotroph, 1 arginine auxotroph, 1 with undetermined zusotrophy
2 serine auxotrophs (serA), 2 early purine auxotrophs (purB), 1 uracil auxotroph (pyrE)
a Mutants classified by auxanography as described previously (13). b Standard auxotroph, able to grow on NB medium but not on minimal medium with or without tetracycline; tetracycline-correctable auxotroph, able to grow on NB
medium with or without tetracycline but grows on minimal E-glucose medium only if tetracycline is provided (specific gene classification based on auxotrophicrequirement and map position; includes strains TT19764 to TT19768); tetracycline-correctable lethal insertion, able to grow on either minimal or NB medium in thepresence of tetracycline [strain TT19197, hlpA1::Tn10d(T-POP)]; tetracycline-enhanced growth rate, growth on NB medium improved by but not requiring the presenceof tetracycline.
suggesting that many genes lack mutant phenotypes and there-
general mechanisms of transcription termination. Currently,
fore were missed by the genetic analysis. The Tn10d(T-POP)
some termination is thought to occur at particular stem-loop
element was used to test this hypothesis and generate single
structures in RNA by mechanisms that are independent of
dedicated protein factors; alternatively, termination can be
Insertions of the Tn10d(T-POP) element in the eut operon
caused by rho factor acting at sites that have proved difficult to
were isolated and tested for correctability of their EutϪ phe-
identify (11, 23, 24). Inspection of the sequences removed by
notype by the addition of tetracycline. Two classes of mutants
the Tn10 deletions reveals differences between the ends of the
were obtained. Insertions in the previously identified eutD, -E,
element. The region downstream of the tetA gene includes a
-A, -B, -C, and -R genes were phenotypically EutϪ with or
stem-loop structure characteristic of rho-independent termina-
without tetracycline. Insertions of Tn10d(T-POP) in the eutS,
tors. Our deletion mutations indicate that this structure is not
-Q, -M, -J, -G, -K, and -L genes showed tetracycline-correctable
a major contributor to transcription termination since only two
aerobic Eut phenotypes as determined with MacConkey etha-
of the five deletions (del24 and del26) remove it. Deletions
indicator medium; previous mutant hunts had
distal to this stem-loop may remove a region necessary for
failed to identify these genes. The new Tn10d(T-POP) inser-
rho-dependent transcription termination which is of major im-
tions demonstrate that these genes were missed because they
portance here. In support of this idea, we have isolated un-
do not encode functions necessary for aerobic metabolism of
linked suppressor mutations that allow tetA-promoted tran-
scripts to extend out of the original Tn10dTet element. Of 20
A secondary use of Tn10d(T-POP) insertions is to select
mutants isolated, 19 carried a rho mutation (data not shown).
in-frame deletions of the identified target region. This has
The sequence removed by the Tn10dTet-internal deletions
been applied to insertions in the eut operon to reinforce the
does not fit well with the features of rho termination sites
conclusion that some genes are nonessential. Tetracycline-cor-
proposed by Alifano et al. (1); they provided evidence that
rectable eut::Tn10d(T-POP) insertion mutants were plated,
rho-dependent terminator regions encode mRNA rich in cy-
without tetracycline, on medium that selects for a Eutϩ phe-
tosine and poor in guanine. The two deletions removing the
notype. Among the revertant clones were deletions which re-
stem-loop show a slightly higher transcription read-through,
moved the T-POP element and target gene sequences; the
suggesting that the stem-loop structure may play a minor role
remaining genes were sufficient for a Eutϩ phenotype. Se-
in addition to a more important rho-mediated termination.
quencing of these revertants showed that the deletions did not
The sequences between the tetR gene and the end of the
disturb the reading frame or else had endpoints near the distal
element agree with general features of rho-dependent termi-
end of genes; these deletions must be essentially nonpolar to
nators in that they show a significant excess of cytosine over
provide the Eutϩ phenotype. Using this deletion procedure, we
guanine. It is interesting to note that only deletions (no point
have confirmed that the eutQ, eutT, eutM, eutN, eutJ, eutG,
mutations) were found to reduce termination of messages. eutH, and eutK genes are nonessential for aerobic utilization of
This is reminiscent of the several mutations (all deletions)
ethanolamine. The implications of eut gene insertions and de-
found to eliminate a rho-dependent termination site within the
letions will be detailed elsewhere (20).
Transcription from the tetA end of Tn10d(T-POP) provides
strong, regulated expression of adjacent genes in a variety of
DISCUSSION
contexts. This is surprising since many adjacent regions would
We describe derivatives of transposon Tn10dTet that allow
be expected to include rho termination sites. It is possible that
regulated, high levels of transcription of chromosomal se-
transcripts emerging from the tetA end of Tn10d(T-POP) may
quences adjacent to the insertion site. The increased transcrip-
be intrinsically resistant to rho termination; alternatively, the
tion results from deletion mutations within the Tn10 element
tetA promoter may be of sufficient strength that commonly
that presumably remove transcription termination signals.
encountered rho sites do not reduce transcription significantly.
These elements (T-POP) should be useful in genetic analysis of
Transcription from the tetR side of Tn10d(T-POP) shows lower
bacteria and in the study of transcription termination.
expression and more variability in its ability to provide tetra-
The use of Tn10d(T-POP) elements permits isolation of
cycline-induced expression of downstream Mud-lac fusions
insertion mutations with conditional (tetracycline-correctable)
than that from the tetA end. Studies on the tet promoters have
phenotypes due to transcriptional fusions between the regu-
shown that mRNA transcripts from the tetR promoter have a
lated tet promoters of the element and a functional chromo-
shorter half-life than transcripts from the tetA promoter (21
somal gene nearby. In principle, these could be formed for any
compared to 41 s) and that the tetR promoter initiates tran-
gene by an insertion between that gene and its promoter.
scription 7 to 11 times less frequently than the tetA promoter
However, since most genes are very close to their promoter,
(10). Insertions at the same his target site but in opposite
such fusions are more common for operons in which an exten-
orientation allowed comparison of transcription levels from
sive target region of dispensable genes is located between the
the tetR and tetA promoters. These results indicate that the tetR
gene providing the phenotype and its normal promoter. We
promoter is 2.5 to 7.5 times less effective than the tetA pro-
describe several auxotrophs, mutants defective for ethanol-
moter (Table 4, rows 1, 2, 3, and 5) at expressing the nearby
amine utilization, and a lethal mutant, whose growth pheno-
lacZ gene. The difference between the earlier promoter studies
types are corrected by tetracycline. Tn10d(T-POP)-generated
and the ability of T-POP promoters to express nearby genes
conditional mutations differ from traditional temperature-sen-
probably reflects factors other than promoter strength; that is,
sitive (i.e., missense) mutations in that nonpermissive condi-
some termination signals may remain within the element or the
tions prevent gene expression instead of altering the native
two transcripts may have different sensitivities to degradation
protein structure; such conditional expression avoids some po-
or termination at sites within sequences adjacent to the inser-
tential complications inherent in the use of temperature-sen-
sitive mutations. The elements described here improve a gen-
Both of the T-POP elements described here have been
eral utility of Tn10dTet elements first pointed out earlier (30).
moved onto FЈ plasmids (TT18797 and TT19196) for use in
Understanding how the described deletions increase tran-
construction of pools of random Tn10d(T-POP) insertions in
scription out of Tn10dTet may provide some insight into the
Salmonella. These plasmids make it possible to introduce T-
POP elements into E. coli by conjugation. Transposition of
a manual for genetic engineering. Cold Spring Harbor Laboratory, Cold
T-POP in E. coli could be selected by transducing the T-POP
element from an FЈ plasmid-bearing strain into an FϪ RecAϪ
14. Faust, L. P., J. A. Conner, D. M. Roof, J. A. Hoch, and B. M. Babior. 1990.
Cloning, sequencing, and expression of the gene encoding the adenosylco-
recipient strain carrying a transposase plasmid (pNK972,
balamin-dependent ethanolamine ammonia-lyase of Salmonella typhi-
pNK2881, or pZT380). This is essentially the method used
murium. J. Biol. Chem. 265:12462–12466.
here to move elements in and out of particular sites in the
15. Halling, S. M., and N. Kleckner. 1982. A symmetrical six-base-pair target
sequence determines Tn10 insertion specificity. Cell 28:155–163.
16. Hillen, W., and C. Berens. 1994. Mechanisms underlying expression of Tn10
encoded tetracycline resistance. Annu. Rev. Microbiol. 48:345–369. ACKNOWLEDGMENTS
17. Hillen, W., and K. Schollmeier. 1983. Nucleotide sequence of the Tn10
encoded tetracycline resistance gene. Nucleic Acids Res. 11:525–539.
We thank Eric Kofoid for help with PCR analysis and DNA se-
18. Hughes, K. T., and J. R. Roth. 1984. Conditionally transposition-defective
derivative of Mud1(Amp Lac). J. Bacteriol. 159:130–137.
This work was supported in part by a grant from the National
19. Kleckner, N., J. Bender, and S. Gottesman. 1991. Uses of transposons with
emphasis on Tn10. Methods Enzymol. 204:139–180.
20. Kofoid, E., C. Rappleye, T. Fazzio, I. Stojiljkvic, D. Sheppard, G. Plunkett, L. Warnick, and J. Roth. Unpublished data. REFERENCES
21. Kofoid, E., and J. R. Roth. Unpublished data.
1. Alifano, P., F. Rivellini, D. Limauro, C. B. Bruni, and M. S. Carlomagno.
22. Raetz, C. R. H. 1996. Bacterial lipopolysaccharides: a remarkable family of
1991. A consensus motif common to all rho-dependent prokaryotic termi-
bioactive macroamphiphiles, p. 1035–1063. In F. C. Neidhardt, R. Curtiss III,
nators. Cell 64:553–563.
J. L. Ingraham, E. C. C. Lin, K. B. Low, Jr., B. Magasanik, W. S. Reznikoff,
2. Benson, N. R., and B. S. Goldman. 1992. Rapid mapping in Salmonella
M. Riley, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and
typhimurium with Mud-P22 prophages. J. Bacteriol. 174:1673–1681. Salmonella: celular and molecular biology, 2nd ed., vol. 1. American Society
3. Berkowitz, D., J. M. Hushon, H. J. Whitfield, J. Roth, and B. N. Ames. 1968.
Procedure for identifying nonsense mutations. J. Bacteriol. 96:215–220.
23. Richardson, J. P. 1993. Transcription termination. Crit. Rev. Biochem. Mol.
4. Berlyn, M. K. B., K. B. Low, and K. E. Rudd. 1996. Linkage map of Esch-
Biol. 28:1–30. erichia coli K-12, Edition 9, p. 1715–1902. In F. C. Neidhardt, (ed.), Esche-
24. Richardson, J. P., and J. Greenblatt. 1996. Control of RNA chain elongation richia coli and Salmonella: cellular and molecular biology, 2nd ed., vol. 2.
and termination, p. 822–848. In F. C. Neidhardt, R. Curtiss III, J. L. Ingra-
American Society for Microbiology, Washington, D.C.
ham, E. C. C. Lin, K. B. Low, Jr., B. Magasanik, W. S. Reznikoff, M. Riley,
5. Bertrand, K. P., K. Postle, L. V. J. Wray, and W. S. Reznikoff. 1984. Con-
M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella:
struction of a single-copy promoter vector and its use in analysis of regula-
cellular and molecular biology, 2nd ed., vol. 1. American Society for Micro-
tion of the transposon Tn10 tetracycline resistance determinant. J. Bacteriol. 158:910–919.
25. Roof, D. M., and J. R. Roth. 1989. Functions required for vitamin B
6. Casadaban, M. J., and S. N. Cohen. 1979. Lactose genes fused to exogenous
dependent ethanolamine utilization in Salmonella typhimurium. J. Bacteriol.
promoters in one step using a Mu-lac bacteriophage: in vivo probe fortranscriptional control sequences. Proc. Natl. Acad. Sci. USA
171:3316–3323. 76:4530–4533.
26. Roof, D. M., and J. R. Roth. 1992. Autogenous regulation of ethanolamine Castilho, B. A., P. Olfson, and M. J. Casadaban. 1984. Plasmid insertion mutagenesis and
utilization by a transcriptional activator of the eut operon in Salmonellalac gene fusion with mini-Mu bacteriophage transposons.
J. Bacteriol. 158:488–495. typhimurium. J. Bacteriol. 174:6634–6643.
8. Chan, R. K., D. Botstein, T. Watanabe, and Y. Ogata. 1972. Specialized
27. Schmieger, H. 1971. A method for detection of phage mutants with altered
transduction of tetracycline resistance by phage P22 in Salmonella typhi-
transducing ability. Mol. Gen. Genet. 110:378–381. murium. II. Properties of a high-frequency-transducing lysate. Virology 50:
28. Schnaitman, C. A., and J. D. Klena. 1993. Genetics of lipopolysaccharide
biosynthesis in enteric bacteria. Microbiol. Rev. 57:655–682.
9. Ciampi, M. S., and J. R. Roth. 1988. Polarity effects in the hisG gene of
29. Stojiljkovic, I. A., A. J. Baumler, and F. Heffron. 1995. Ethanolamine utili-
Salmonella require a site within the coding sequence. Genetics 118:193–202.
zation in Salmonella typhimurium: nucleotide sequence, protein expression
10. Daniels, D. W., and K. P. Bertrand. 1985. Promoter mutations affecting
and mutational analysis of the cchA cchB eutE eutJ eutG eutH gene cluster.
divergent transcription in the Tn10 tetracycline resistance determinnt.
J. Bacteriol. 177:1357–1366.
J. Mol. Biol. 184:599–610.
30. Takiff, H. E., T. Baker, T. Copeland, S. Chen, and D. L. Court. 1992.
11. Das, A. 1993. Control of transcription termination by RNA-binding proteins.
Locating essential Eschericia coli genes by using mini-Tn10 transposons: the
Annu. Rev. Biochem. 62:893–930. pdxJ operon. J. Bacteriol. 174:1544–1553.
12. Davis, R. W., D. Botstein, and J. R. Roth. 1980. Advanced bacterial genetics.
31. Way, J. C., M. A. Davis, D. Morisato, D. E. Roberts, and N. Kleckner. 1984.
Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
New Tn10 derivatives for transposon mutagenesis and for construction of
13. Davis, R. W., D. Botstein, and J. R. Roth. 1980. Advanced bacterial genetics: lacZ operon fusions by transposition. Gene 32:369–379.
Zoopharmacognosy and Your Animals Have you ever noticed your horse, dog or cat eating a certain plant in your garden or out in the fields? Well, if you have, you may be witnessing Zoopharmacognosy. This is a term coined by Dr. Eloy Rodriguez, a biochemist and professor at Cornell University. Zoopharmacognosy refers to the process by which animals self-medicate. In this process, animals select
PREDICTING THE EFFECTIVENESS OF HYDROXYUREA IN INDIVIDUAL SICKLE CELL ANEMIA PATIENTS Homayoun Valafar, Faramarz Valafar, Alan Darvill and Peter Albersheim, Complex Carbohydrate Research Center and the Department of Biochemistry and Molecular Biology, University of Georgia, 220 Riverbend Road, Athens, GA 30602 Abdullah Kutlar, Kristy F. Woods, and John Hardin, Department of Medicin
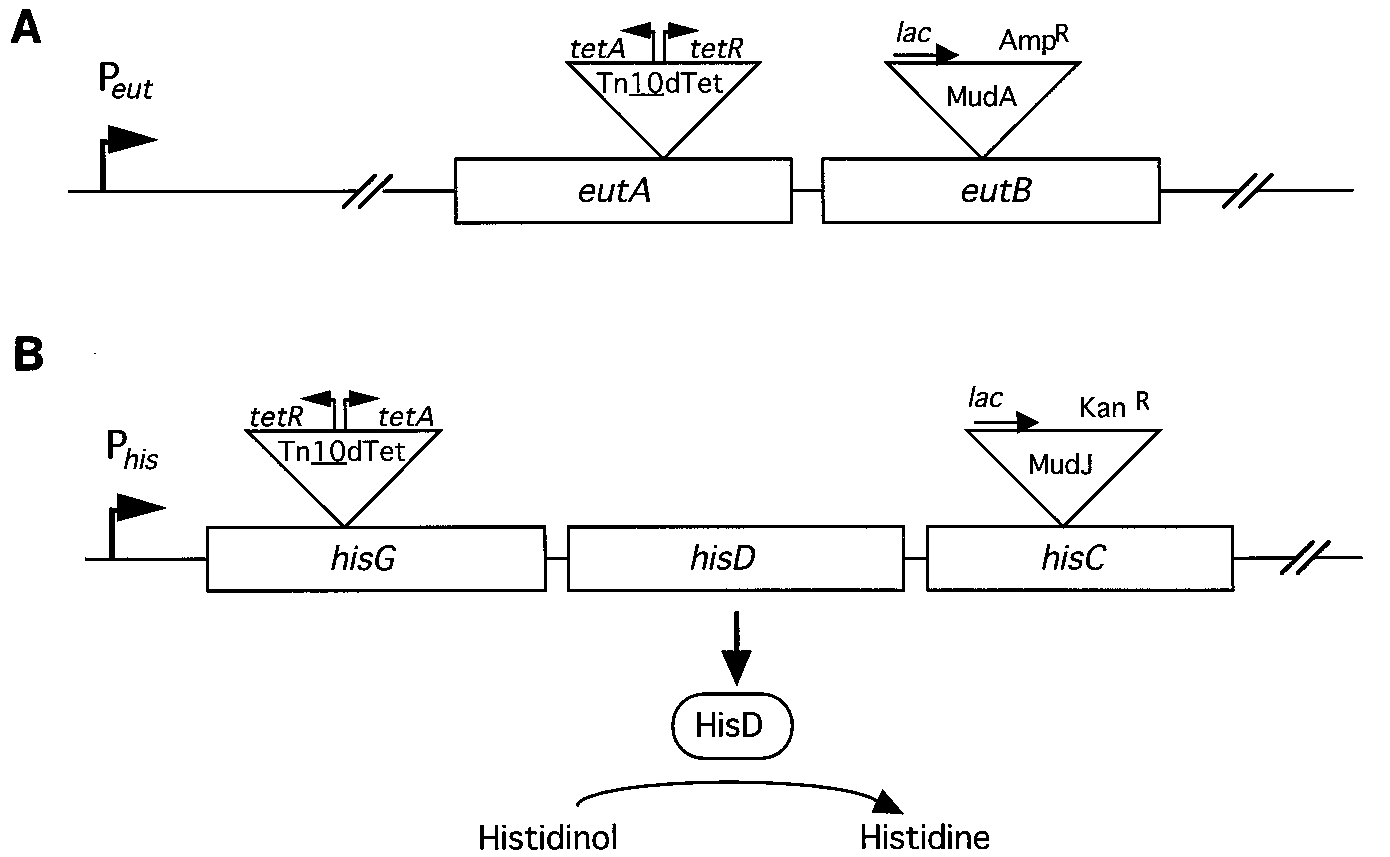 FIG. 1. Selection systems for modification of Tn10dTet. These abbreviated maps show the positions of Tn10dTet and MudJ insertions within the ethanolamine (eut)
(A) and histidine (his) (B) operons of the parent strains. The eutA and eutB genes are the 11th and 12th genes in the ethanolamine operon. The hisG, -D, and -C genesare the first three genes in the histidine operon. The hisG::Tn10dTet insertion is at a hot spot for Tn10 insertion.
FIG. 1. Selection systems for modification of Tn10dTet. These abbreviated maps show the positions of Tn10dTet and MudJ insertions within the ethanolamine (eut)
(A) and histidine (his) (B) operons of the parent strains. The eutA and eutB genes are the 11th and 12th genes in the ethanolamine operon. The hisG, -D, and -C genesare the first three genes in the histidine operon. The hisG::Tn10dTet insertion is at a hot spot for Tn10 insertion.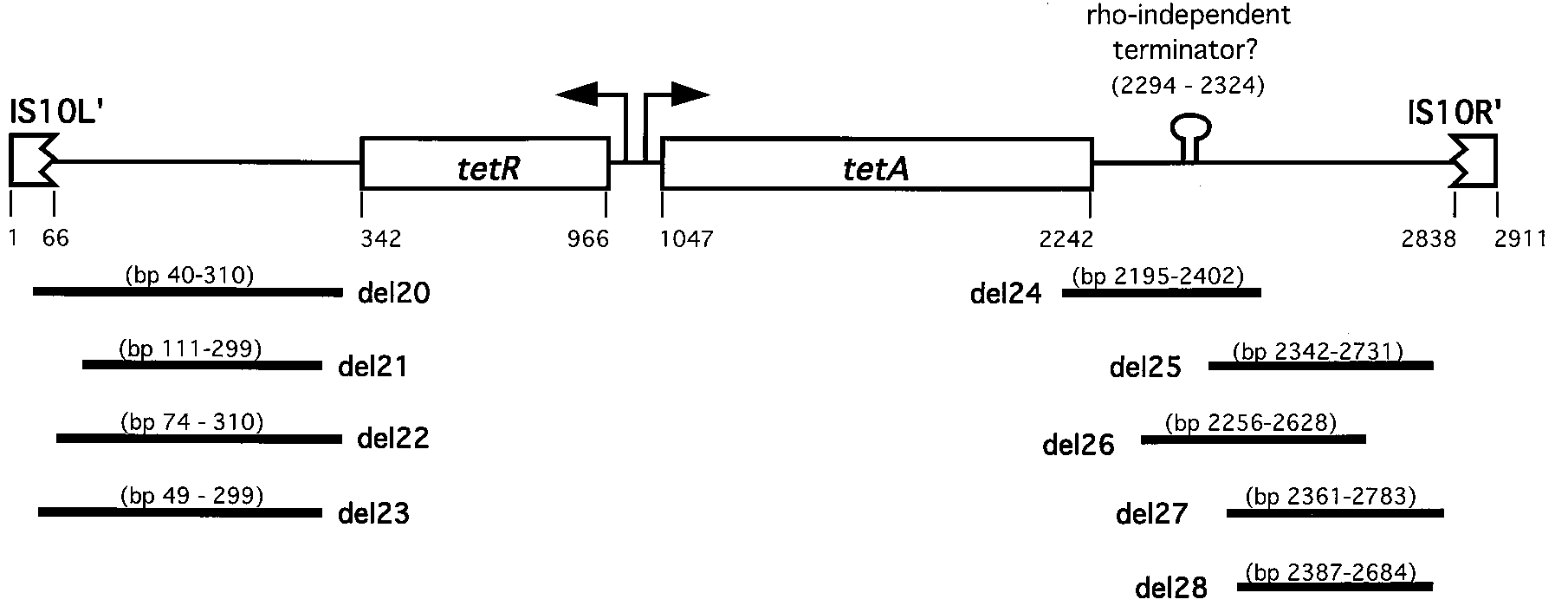 FIG. 2. Position of deletion mutations within Tn10dTet. Deletions at the tetR end (left) were selected by use of the insertions in the ethanolamine operon; those
at the tetA end (right) were selected by use of the insertions in the histidine operon. Numbers indicate the positions of each feature in the DNA base sequence of theparent transposon, Tn10dTet.
FIG. 2. Position of deletion mutations within Tn10dTet. Deletions at the tetR end (left) were selected by use of the insertions in the ethanolamine operon; those
at the tetA end (right) were selected by use of the insertions in the histidine operon. Numbers indicate the positions of each feature in the DNA base sequence of theparent transposon, Tn10dTet.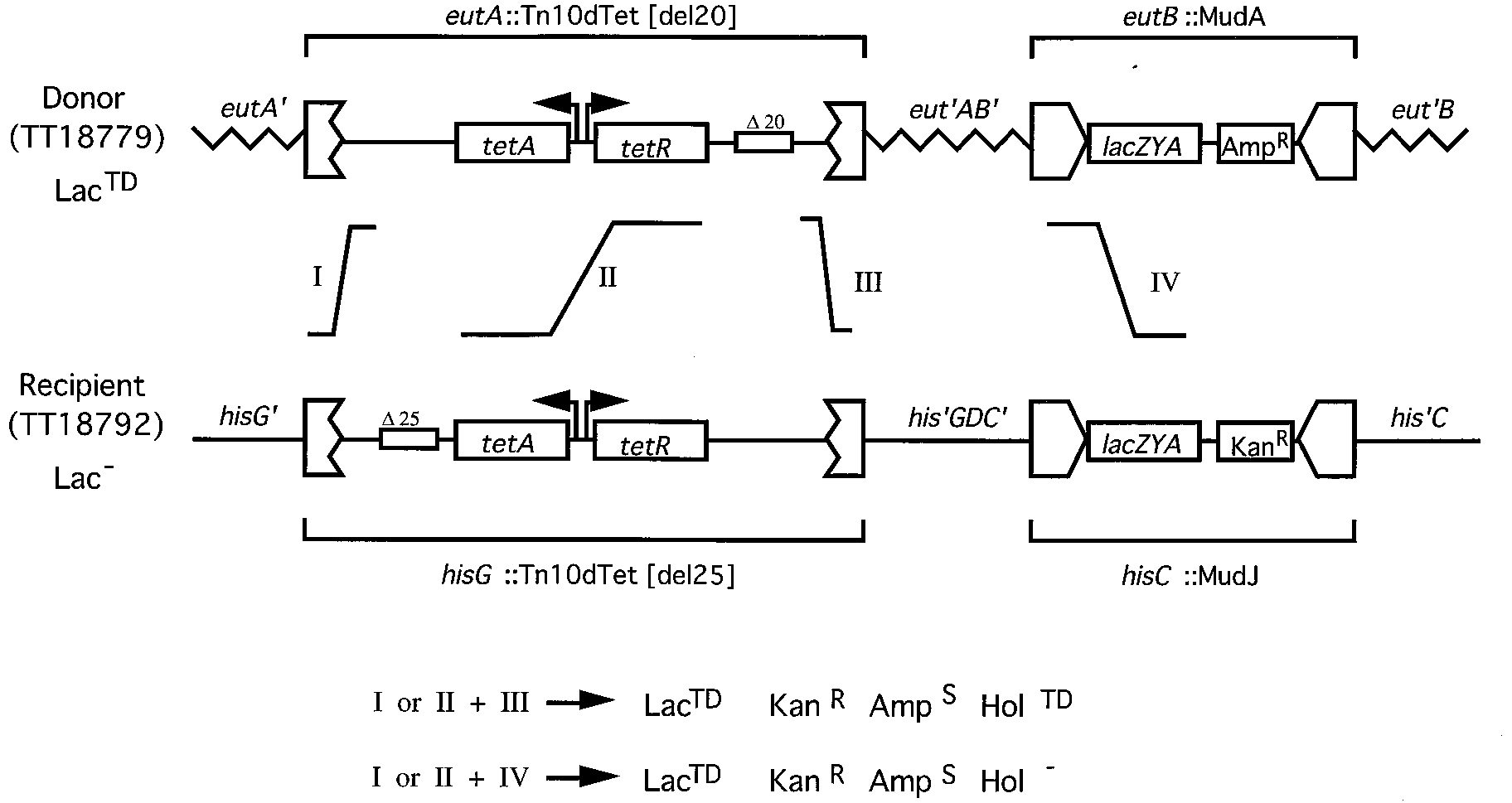
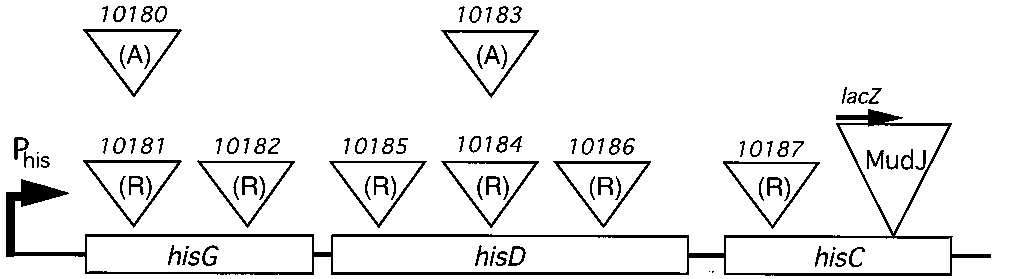 FIG. 3. Construction of a transposon with two deletion mutations. The diagrammed cross was used to construct T-POP, the double deletion mutant Tn10dTet
(del25, del20). The recipient carries mutation del25 but is oriented such that the end needed for expression of the MudJ lac genes has no deletion; this strain isphenotypically LacϪ. The donor strain has mutation del20 at the end nearest the MudA-lac operon. Therefore, this strain is phenotypically LacTD. In the cross, selectionis made for LacTD recombinants; this demands inheritance of the donor mutation del20 (events allowing this are indicated by roman numerals). The donor elementscannot be inherited at the recipient eut locus because it carries a large eut deletion mutation. All recombination events must occur within the two transposons sequencesshared by donor and recipient.
FIG. 3. Construction of a transposon with two deletion mutations. The diagrammed cross was used to construct T-POP, the double deletion mutant Tn10dTet
(del25, del20). The recipient carries mutation del25 but is oriented such that the end needed for expression of the MudJ lac genes has no deletion; this strain isphenotypically LacϪ. The donor strain has mutation del20 at the end nearest the MudA-lac operon. Therefore, this strain is phenotypically LacTD. In the cross, selectionis made for LacTD recombinants; this demands inheritance of the donor mutation del20 (events allowing this are indicated by roman numerals). The donor elementscannot be inherited at the recipient eut locus because it carries a large eut deletion mutation. All recombination events must occur within the two transposons sequencesshared by donor and recipient.