Die Struktur von Tadalafil erlaubt eine selektive Bindung an die Bindungsstelle der PDE5 und minimiert gleichzeitig die Interaktion mit PDE6, was visuelle Nebenwirkungen einschränkt. Seine Verteilung im Organismus erfolgt breit, wobei das Verteilungsvolumen etwa 63 Liter beträgt. Über 90 % des Wirkstoffs sind an Plasmaproteine gebunden. Die Wirkung bleibt unabhängig von der Nahrungsaufnahme konstant. Der Abbauweg über CYP3A4 kann durch Hemmer wie Ritonavir oder Ketoconazol verlangsamt werden, was die Plasmakonzentrationen deutlich erhöht. In diesem Kontext wird cialis 20mg preis häufig in Bezug auf pharmakokinetische Wechselwirkungen erwähnt.
071-82 vogel.qxd
Edgar H. Vogel1, María E. Castro2, Paola A. Solar2, and Fabián A. Soto1
1University of Talca, School of Psychology, Casilla No 747, Talca, Chile;2University of Mar, Laboratory of Neurosciences, Carmen 446, Cerro Placeres,
Abstract. The goal of this study was to define conditions under which
conditioned immunosuppression may be observed reliably. In three
experiments, rats were exposed to a gustatory conditioned stimulus (CS) paired
with cyclophosphamide (US), which induces immunosuppression and malaise.
In Experiment 1, a single pairing of the CS with low, medium, or high doses
of cyclophosphamide in separate groups produced no reliable conditioned
immunosuppression even though conditioned taste aversion was observed
in groups trained with high and medium doses of CY. Experiment 2 replicated
the lack of effect following a single pairing of the CS with the medium dose
of cyclophosphamide but demonstrated that three pairings are sufficient
to induce conditioned immunosuppression. Experiment 3 demonstrated that
significant immunosuppression is observable following a single CSUS
pairing if the CS is presented in compound with a previously nonreinforced CS
during training, an effect reminiscent of supernormal conditioning. These
findings indicate that conditioned immunosuppression effects can be enhanced
in magnitude through the use of certain procedural techniques.
Key words: conditioned immunomodulation, conditioned inmunosupression,
Pavlovian conditioning, psychoneuroimmunology
detectable only when the CS is accompanied by a sub-
threshold booster dose of the antigen US (Ader et al.
Over 30 years of research has established that 1993).
immune responses can be modulated by Pavlovian
Small and sometimes inconsistent conditioned
conditioning procedures. In these experiments, an immunosuppression effects have been observed as
immunologically neutral conditioned stimulus (CS) is well. The simplest type of conditioned immunosup-
paired with an immunologically active unconditioned pression experiment measures humoral immunity after
stimulus (US) and thereby comes to control a condi- pairing a saccharin solution with CY. The experimen-
tioned response (CR) that is similar to the uncondi- tal design typically consists of three phases. In the
tioned response controlled by the US. Initially, this conditioning phase, a conditioned group of rats is
phenomenon was studied using immunosuppressive exposed to a single pairing of a saccharin solution (CS)
drug USs, such as cyclophosphamide (CY) or and an IP injection of CY (US). In the testing phase,
cyclosporin A, paired with gustatory CSs, in what has which occurs several days later, conditioning is evalu-
been called the conditioned inmunosupression para- ated by exposing the rats to the CS in conjunction with
digm (Ader and Cohen 2001). In this paradigm, the an immunological challenge consisting of antigen
conditioned effects consist of a decreased antibody inoculation (SRBC). In the third phase, immune assays
response to certain antigens (typically, sheep red blood are conducted to quantify the immune reaction to the
cells or SRBC) in the presence of the gustatory CS. In antigen. Antibody titers in the conditioned group are
subsequent studies using similar CSs and USs, other compared to those in several control groups, which can
conditioned immunological responses have been be classified into two categories: the placebo group
observed, such as decreased graft-versus-host respons- and the nonconditioned groups. In the placebo group,
es (Bovbjerg et al. 1984) and reduction in natural killer the animals receive a saline injection instead of CY and
cell activity (OReilly and Exon 1986). More recently, are expected to exhibit no immunosuppression at the
researchers have become interested in the possibility of time of testing (neither conditioned, due to the CS, nor
conditioned immunoenhancement through the use of unconditioned, due to residual effects of CY). In the
immune system-potentiating drugs (Solvason et al. nonconditioned groups, the animals receive the same
1991) and antigen USs (Ader et al. 1993, Alvarez- dose of CY as does the conditioned group, but no CS-
Borda et al. 1995, Gorczynski et al. 1982, Madden et US pairings occur. This can be done in any of several
ways, including presenting the US but not the CS dur-
The finding that the immune system is conditionable ing the conditioning phase (US only group); omitting
has stimulated a respectable volume of research, con- the CS during testing in a conditioned group (CSo
tributing greatly to the development of an emergent group); and/or presenting the CS and the US in an
discipline known as psychoneuroimmunology. The explicitly unpaired fashion (unpaired group). The non-
corpus of the empirical data and its clinical implica- conditioned groups are expected to exhibit no
tions have been reviewed periodically by Ader, Cohen, immunosuppression due to conditioning and little, if
and their colleagues (Ader 1981, 2003, Ader and any, immunosuppression due to the residual effects of
Cohen 1991, 2001, Cohen et al. 1994) as well as CY. Thus, to probe the effects of conditioning, it is nec-
others (e.g., Dunn 1989, Exton et al. 2000, essary to demonstrate that the conditioned group is
Hucklebridge 2002, Kusnecov et al. 1989, Stockhorst more immunosuppressed than is any of the noncondi-
and Klosterhalfen 2005). There is considerable agree- tioned groups. Since CY also produces malaise, the
ment among the reviewers as to the reality of the con- effectiveness of the CS/US pairings can be further con-
ditioned immunomodulation phenomenon, but there is firmed by observing a decreased consumption of sac-
also some concern with respect to the reliability and charine in the conditioned group as compared to the
size of the effects. For instance, Ader (2003) discussed control groups (i.e., conditioned taste aversion).
some of the failures to replicate the finding of condi-
There are a few studies on humoral conditioned
tioned immunoenhancement, and concluded that these immunosuppression similar to the one described
studies may have failed to detect what is often a small above. For instance, in the first report by Ader and
effect. Support for this idea comes from the finding Cohen (1975), the results were in the expected direc-
that conditioned immunomodulation sometimes is tion; namely, the paired group exhibited the lowest
Enhancement of conditioned inmunosuppression 73
antibody titers, the placebo group exhibited the highest The rationale of this procedure is that given that CS B
antibody titers, and the nonconditioned groups were by itself developed a negative association with the US
intermediate. Subsequently Ader and coauthors (1982) and that the AB compound is followed by the US, the
reported similar findings in a study that included con- amount of association developed by A should be
ditioned, placebo, and CSo groups. Similar experi- greater than normal in order to counteract the negative
ments were conducted by Rogers and others (1976) expectation created by the presence of B.
and Wayner and others (1978) and, although the data
were in the same direction as in previous studies, the
Collectively, these experiments on humoral
immunosuppression support the idea that immune
responses can be conditioned, since they found approx-
One hundred eighty-two male albino rats (purchased
imately the same pattern of results. However, taken from Pontificia Universidad Católica de Chile breed-
individually, none of these experiments provides con- ing colony, Santiago, Chile) weighing 200350 g, were
clusive evidence of a robust conditioned immunosup- housed in individual cages with free access to food.
pressive response. Furthermore, the results of some of Mounted on each cage was a holder that could support
these studies suggested that in order to see condition- a 100-ml plastic syringe adapted with a glass-drinking
ing, the experimental protocol must include at least spout at the bottom. All fluids were delivered in these
syringes. All experimental treatments took place in the
An additional source of uncertainty comes from individual home cages. Different animals were used
a few studies that report opposite conditioned effects; for each experiment (n=94 for Experiment 1, n=44 for
i.e., immunoenhancement instead of immunosuppres- Experiment 2, and n=44 for Experiment 3). Rats in
sion following training with an immunosuppressive experiments 1 and 2 had restricted access to water at
drug (e.g., Krank and McQueen 1988, McQueen and some stages of the experiment whereas rats in
Experiment 3 were kept on an ad libitum regimen of
The present set of experiments was designed to water throughout the experiment.
explore the possibility of obtaining more reliable and
All experimental protocols were approved by
robust conditioned immunosuppression effects using Universidad de Talca Ethics Committee and CONYC-
a classic experimental procedure in which rats are IT(Comisión Nacional de Investigación Científica y
exposed to a gustatory CS and cyclophosphamide US. Tecnológica de Chile).
Experiment 1 was designed to reproduce the findings
of Ader and Cohen (1975) using saccharin and three
different intensities (low, medium and high doses) of
CY. Having obtained no evidence of conditioned
CONDITIONING PROTOCOL OF EXPERIMENT 1
immunosuppression in this experiment despite of
robust conditioned taste aversion, we next employed
Experiment 1 was designed to evaluate conditioned
two further enhancement strategies. In Experiment 2, immunosuppression after training separate groups of
a reliable conditioned immunosuppression effect was animals with three different doses of CY (low, medium
observed after increasing the number of saccharin-CY and high). The experiment was conducted in two repli-
pairings from one to three. In Experiment 3, reliable cations, each including 47 animals. The two replica-
conditioned immunosuppression was obtained follow- tions were identical except as noted below. One week
ing a single saccharin-CY pairing when saccharin was after arrival, the rats were adapted over a 7-day period
presented in compound with lemon juice, which had to 15 minutes of access to water per day. After adapta-
been trained to signal the absence of CY in a nonrein- tion, the rats in each replication were matched into the
forced pre-exposure phase. This later strategy resem- 7 groups based on their water intake and body weight.
bles the so called supernormal conditioning proce-
Table I outlines the experimental treatments for each
dure, which refers to the enhancement of the learning group. On the day following the last day of adaptation
that accrues to a given CS, A, after being compounded (day 1), animals in the groups unpaired-high, unpaired-
with an inhibitory CS, B, during the conditioning trials. medium and unpaired-low were exposed to a novel
taste by replacing the plain water with a 0.1% sodium
Thereafter all animals were returned to daily 15-min
saccharin (SAC) solution during the 15-min drinking access to plain water, except on the test day. The test
period. On this day, all other groups received the regu- was conducted on day 5 (replication 1) or day 12 (repli-
lar 15-min access to plain water. The next day (day 2) cation 2) and consisted of the replacement of plain
was the conditioning day. Animals in groups unpaired- water with the saccharin solution in all groups.
high, unpaired-medium and unpaired-low received Immediately after finishing the 15-minute drinking
plain water, and all other groups received flavored period, all animals were immunized with i.p. injections
water. Thirty minutes after the drinking period, ani- of 2 ml/kg of a 1% thrice-washed suspension of SRBC.
mals in the vehicle group received an intraperitoneal Seven days after immunization (day 11 for replication
(i.p.) injection of 1 ml of physiological saline, and all 1 and day 18 for replication 2), blood was drawn by car-
other groups received an i.p. injection of CY. The dose diac puncture performed in anesthetized animals. The
of CY was of 75 mg/kg in groups paired-high and blood was centrifuged and the serum was collected and
unpaired-high, of 50 mg/kg in groups paired-medium inactivated at 57°C for 30 min prior to determination of
and unpaired-medium, and of 25 mg/kg in groups antibody titers by the hemagglutinating microtiter
method described by Ader and Cohen (1975).
Enhancement of conditioned inmunosuppression 75
CONDITIONING PROTOCOL OF EXPERIMENT 2
strength when presented in compound with taste B as
compared to when presented alone. That is, similar to
Experiment 2 was conducted to test whether the the supernormal conditioning phenomenon, taste A
conditioned immunosuppression effect can be aug- might be assigned greater than normal significance as
mented by increasing the number of CSUS pairings. a function of its correctly signaling an outcome (US
The conditioning and test protocols were similar to delivery) different from that signaled by taste B (no US
those of the second replication of Experiment 1, except delivery). The major features of this experiment are
that only a medium dose of CY was used (50 mg/kg). outlined in Table III. The animals were divided into 4
Table II outlines the different treatments. Animals in groups that were treated differentially in two experi-
groups paired-one trial and unpaired-one trial received mental phases, the pre-exposure phase and the condi-
a single dose of CY (day 13), whereas animals in tioning phase. Group superconditioning was exposed
groups paired-three trials and unpaired-three trials to taste B during the pre-exposure phase and to the AB
received three doses of CY (days 3, 8 and 13). Animals compound followed by CY during the conditioning
in groups paired-one trial and paired-three trials were phase. Group Paired also was exposed to taste B dur-
exposed to the saccharin solution 30 minutes before ing the pre-exposition phase, but during conditioning A
being injected with CY, whereas animals in groups was presented in isolation (i.e., not as part of an AB
unpaired-one trial and unpaired-three trials were compound) and paired with CY. It was expected that
exposed to saccharin two days before the injection of the pre-exposure to taste B would not have any influ-
CY. Similar to replication 2 of Experiment 1, immu- ence over conditioning to the qualitatively different
nization occurred 10 days after the last day of condi- taste A in this group. Two nonconditioned control
tioning and blood samples were drawn 7 days after groups were included in the design. Animals in the
unpaired control group were exposed to taste B during
all trials of the pre-exposure and conditioning phases,
CONDITIONING PROTOCOL OF EXPERIMENT 3
except for pre-exposure trial number 3, in which they
were exposed to taste A approximately 24 hours before
Experiment 3 was conducted to test whether the the administration of CY. This arrangement constituted
conditioned immunosuppression effect can be aug- an unpaired presentation of taste A and CY, such that
mented by presenting a target CS, A, in compound with no conditioning should occur to taste A. Animals in the
another CS, B, that previously had been trained to sig- US control group were exposed to no flavor at any
nal the absence of the US. The rationale was that ani- point in the experiment, but received a CY injection
mals should learn that taste B was not followed by any during the conditioning phase. This new control group
consequence during the pre-exposure phase, and as was included to have a measure of the unconditioned
a result, taste A might acquire more associative residual effect of CY administration upon the immune
response, without the influence of exposure to the con- tions. As can be seen in the table, there were no major
differences between the groups in the first exposure
The pre-exposure and conditioning phases lasted to the saccharin solution. This result was confirmed
three days, each of which was divided into morning by an ANOVA in which no significant main effect of
and evening sessions (at about 8:00 A.M. and 8:00 P.M. group was noted in the conditioning session
respectively). The four pre-exposure sessions occurred (F =1.583, P=0.162, partial h2=0.098). Group dif-
in the afternoon session of day 1, in the morning and ferences arose during the testing day and were con-
afternoon session of day 2, and in the morning session firmed by a significant main effect of group
of day 3. Conditioning took place in the afternoon ses- (F =4.718, P<0.001, partial h2=0.245). As can be
sion of day 3, where all animals received a 50 mg/kg seen in the table, the unpaired (high, medium, and
of CY. Unlike experiments 1 and 2, in this experiment low) and vehicle groups did not show evidence of
the animals were exposed to the flavors by forcing taste aversion, while groups paired-high and paired-
them to drink 1 ml of fluid by pipette. Immunization medium showed a large decrement in their preference
occurred 4 days after the last day of conditioning (day ratios. Contrary to expectations, the paired-low group
7). During immunization, all animals were exposed to did not show evidence of CTA. Planned contrasts
the target taste A. Blood samples were drawn 7 days confirmed the reliability of these observations since
group paired-high differed significantly from groups
unpaired high and vehicle (Ps=0.011 and 0.051; par-
tial h2s=0.072 and 0.043, respectively), and group
paired-medium differed significantly from groups
In Experiments 1 and 2, conditioned taste aversion unpaired-medium and vehicle (Ps=0.001 and 0.005;
(CTA) was assessed through a preference ratio calcu- partial h2s=0.138 and 0.088, respectively). Finally,
lated for each animal by dividing their water intake group paired-low did not differ significantly from
during the test by the mean water intake on the two group unpaired-low or from group vehicle (Ps=0.427
immediately preceding days. A preference ratio of less and 0.791; partial h2s=0.007 and 0.001, respectively).
than 1 suggests that an aversion was developed to the These results indicate that there was an association
taste, whereas a value equal to or higher than 1 indi- between saccharine and CY at least in groups paired-
cates the absence of aversion. In Experiment 3, CTA high and paired-medium.
was not examined. Conditioned immunomodulation
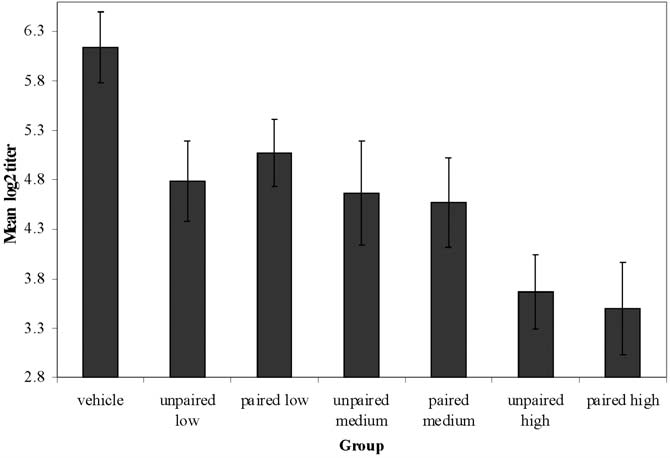
Figure 1 presents the mean antibody titers in the
was assessed through the antibody titers recorded as 7 groups. As expected, group vehicle showed the high-
log2 reciprocals of the end point dilutions in est immune response, followed by the two groups that
were trained with a low dose of CY, then by the two
One-way ANOVAs were conducted to test the main groups trained with an intermediate dose, and finally by
effect of group in each experiment. Planned compar- the two groups trained with the high dose. Since all
isons using the Fisher PLSD technique were employed
in order to test for relevant differences among specific
groups. All statistical differences were considered sig-
nificant at the 0.05 level. Effect sizes were reported as
Mean preferences ratios and standard error of the mean
(SEM) in the training and test trials of Experiment 1
Experiment 1: Effects of pairing a saccharin
solution with three different doses of CY
Table IV presents the preference ratios for the
seven groups in the training and test trials. Because
equivalent results were obtained between replica-
tions, the data were collapsed across the two replica-
Enhancement of conditioned inmunosuppression 77
The main effect of group was explored further
through nine planned contrasts. The first three contrasts
indicate that there was no reliable difference between
groups paired-high and unpaired-high (P=0.784, partial
h2=0.001); between groups paired-medium and
unpaired-medium (P=0.876, partial h2=0.000); or
between groups paired-low and unpaired-low (P=0.626,
partial h2=0.003). On the other hand, the mean antibody
titers of group vehicle was significantly greater than that
of every other group (Ps<0.07, partial h2s>0.077), indi-
cating a residual effect of CY. Thus, contrary to what
was observed with CTA, there was no evidence of con-
ditioned immunomodulation in this experiment.
Fig. 1. Mean hemagglutination titers (± standard error of the
Experiment 2: Effect of increasing the number
groups that were exposed to CY exhibited some degree
of immunosuppression, the critical comparisons to
Evidence of CTA was again found in this experi-
demonstrate conditioning are between the respective ment. Table V presents the mean preference ratios for
paired and unpaired groups. As the figure shows, these all groups in the testing trials. As expected, there were
differences were very minor. The data were explored no major differences between the groups on the first
with a 2 (replication) × 7 (group: paired-high; unpaired- trial, as confirmed by an ANOVA indicating no reliable
high; paired-medium; unpaired-medium; paired-low; effect of group in this trial (F =0.789, P=0.507, par-
unpaired-low; vehicle) ANOVA, which indicated a reli- tial h2=0.056). Group differences arose during the sec-
able main effect of replication (F =5.913, P=0.017, ond testing day (F =10.325, P<0.001, partial
partial h2=0.069) and group (F =5.206, P<0.001, par- h2=0.436). As can be seen in the table, group paired-
tial h2=0.281), but no reliable interaction between three trials showed an important decrement of water
group and replication (F =1.791, P=0.111, partial intake in trial 2 and the lowest preference ratios among
h2=0.118). The main effect of replication is due to the the groups. Planned comparisons confirm the reliabili-
fact that the antibody titers of replication 1 were lower ty of this CTA, indicating that group paired-three trials
than those in replication 2, which is explained by the presented a preference ratio significantly lower than
difference in the interval between CY administration that of group unpaired-three trials (P<0.001, partial
and immunization in the two replications (4 versus 10 h2=0.383) and that groups paired-one trial and
days). Since there was no interaction between group unpaired-one trial did not differ significantly
and replication, no further analysis was pursued with (P=0.988, partial h2=0.000). Reliable differences
the later variable and the data are presented collapsed between groups were again found during trial 3
(F =6.526, P=0.001, partial h2=0.329), with planned
Mean preferences ratios and standard error of the mean (SEM) in the four testing trials of Experiment 2
tive unpaired group. This latter analysis revealed a reli-
able conditioning effect in group paired-three trials,
whose mean immune response was significantly lower
than the unpaired-three trials group (P=0.038, partial
h2=0.103). Groups paired one-trial and unpaired one-
trial did not differ reliably (P=0.648, partial h2=0.005).
In summary, Experiment 2, like Experiment 1,
showed an important dose-dependent residual effect of
CY and no reliable conditioned immunosuppressive
effects after training with a single dose of CY.
However, this experiment showed that three CSUS
pairings may be more effective in producing reliable
Experiment 3: Effects of training a target CS
Fig. 2. Mean hemagglutination titers (± standard error of the
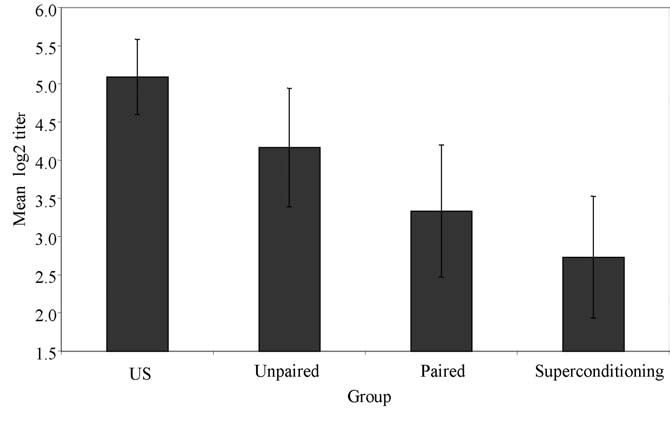
Figure 3 presents the mean antibody titers for all
tests indicating a reliable difference between groups groups in this experiment. The highest mean was
paired-three trials and unpaired-three trials (P<0.001, exhibited by the US control group, followed by the
partial h2=0.303) and no reliable difference between unpaired group, the paired group and, finally, the
groups paired-one trial and unpaired-one trial superconditioning group. Given that the mean anti-
(P=0.187, partial h2=0.043). In trial 4, which tested body titers of the superconditioning group was lower
learning after three CSUS parings in paired-three tri- than that of the paired group, it seems that the super-
als group and learning after a single paring in the conditioning procedure was effective in enhancing
paired one-trial group, the ANOVA indicated reliable learning of a conditioned immunosuppressive response
differences among the groups (F =20.954, P<0.001, to the target taste. Although the simple main effect of
partial h2=0.611). In this last trial, planned compar- group was not reliable (F =2.004, P=0.129, partial
isons showed that the two conditioned groups (paired- h2=0.131), planned comparisons showed a reliable dif-
three trials and paired-one trial) differed significantly ference between group superconditioning and the
from their respective unconditioned control groups pooled control groups (P=0.037, partial h2=0.104),
(unpaired-three trials and unpaired-one trial; Ps=0.000 whereas group paired, as in all the preceding experi-
and 0.010; partial h2s=0.580 and 0.153, respectively). ments, did not differ reliably from the control groups
In summary, CTA was apparent in the two conditioned (P=0.162, partial h2=0.048). The differences between
groups and seems to be stronger as the number of CS-
Figure 2 depicts the results of the immune assays for
the four groups. As can be seen, there were clear dif-
ferences among the groups that were confirmed by
a reliable main effect of group (F =10.271, P<0.001,
partial h2=0.435). As expected, the number of CY
innoculations had a major impact on the immune
response, as confirmed by planned contrasts comparing
the two groups that received a single dose of CY (i.e.,
paired-one trial and unpaired-one trial) with the groups
that received three doses (i.e., paired-three trials and
unpaired-three trials; P<0.001, partial h2=0.611). The
possibility of conditioning was examined by planned Fig. 3. Mean hemagglutination titers (± standard error of the
contrasts comparing each paired group with its respec- mean) in Experiment 3
Enhancement of conditioned inmunosuppression 79
groups superconditioning and paired, and between
Experiment 3 demonstrated that it is possible to
both control groups, were not reliable according to enhance conditioned immunosuppression without
planned comparisons (Ps=0.567 and 0.362; partial appealing to several doses of the drug by means of
h2s=0.08 and 0.021, respectively). Thus, although con- a procedure that resembles supernormal conditioning.
ditioning a target CS in compound with a pre-exposed In the standard supernormal conditioning procedure,
CS did not produce an immunosuppressive response learning to a target stimulus is enhanced when it is rein-
greater than that observed by normal CS/US pairings, forced in compound with an inhibitory stimulus
it was large enough to support a reliable conditioned (Rescorla 1971, 2004, Wagner 1971, Williams and
immunosuppression effect whereas the paired proce- McDevitt 2002). Considering that the training of a con-
ditioned inhibitor normally requires several presenta-
tions of the US, and that one of our goals was to avoid
the chronic effects of repeated drug administration, we
attempted to obtain a superconditioning-like effect
Taken together, Experiments 13 suggest that the without a conditioned inhibitor. Hence, we trained a CS
conditioning procedure based upon a single pairing of to signal the absence of reinforcement by pre-exposing
a gustatory CS with CY is effective in producing it without any consequence before conditioning. Then
robust CTA but does not lead to a comparable condi- we presented a compound of this pre-exposed CS and
tioned immunosuppression. In all three experiments, a target CS, followed by CY. This procedure was suffi-
animals that received a single pairing of CY with sac- cient to obtain a significant conditioned response that
charin presented an immune response that was not sig- was not seen with the regular CS/US pairing.
nificantly lower than that of the animals that received
A common explanation of supernormal conditioning
the same stimuli in an unpaired fashion.
is that, during compound conditioning, the inhibitory
It should be pointed out that in the present experi- stimulus creates an expectation of non-reinforcement
ments the immunosuppressive response was tested that is violated when reinforcement is presented, aug-
using a single presentation of the gustatory CS menting excitatory conditioning (e.g., Rescorla and
(i.e., a single test trial), whereas previous research Wagner 1972). Although the presentation of an inhibitory
showing evidence of conditioned immunosuppression stimulus is the most common way to produce supernor-
(e.g., Ader and Cohen 1975, Ader et al. 1982, Rogers et mal conditioning to a target CS, there are other ways to
al. 1976) used a slightly different conditioning proce- obtain the effect. For example, Dickinson (1977) found
dure, in which two test trials were included. Therefore, that fear conditioning to a target CS is enhanced when it
it is possible that the negative results reported here may is reinforced (i.e., paired with shock) in compound with
be the result of a small-sized effect that could be a stimulus previously paired with food. An interpretation
enhanced by the use of several test trials. The results of of this result is that the presence of the CS previously
the present experiments suggest other factors that could paired with food during compound conditioning gener-
enhance the conditioned immunosuppression effect.
ates an expectation of an appetitive outcome, which is
The results from Experiment 1 indicate that manip- violated when the shock, an aversive outcome, is pre-
ulating the salience of the US (i.e., the CY dose) may sented. In this case, what seems to be the critical condi-
not be an effective method to uncover potential condi- tion to produce supernormal conditioning is the presen-
tioned effects in this paradigm. In Experiment 2, rather tation of a US that is contrary to the expectations of rein-
than increasing the salience of the US, we increased forcement based on previous learning. It seems that more
the number of CSUS trials. This was, in fact, a much conditioning occurs whenever reinforcement is particu-
more effective method that has been used by others to larly surprising for the organism. The results of
demonstrate the clinical impact of conditioning. For Experiment 3 are consistent with this view.
instance, Ader and Cohen (1982) reported that three
It should be noticed that compounding a target CS
saccharin/CY pairings delayed the development with a pre-exposed stimulus does not always lead to an
of lupus in mice. Of course, increasing the number of augmentation of conditioning. For instance, Navarro
conditioning trials implies prolonged exposure to the and others (1989) compared fear conditioning to a target
immunosuppressive drug and its chronic effects upon stimulus after training it either in compound with
the immune response, which may be less than ideal. a pre-exposed CS (supernormal condition) or with
a novel CS (normal condition) and found no differences Ader R, Cohen N (1991) The influence of conditioning on
between the two conditions. The results of our
immune responses. In: Psychoneuroimmunology (Ader
Experiment 3 are similar to those reported by Navarro
R, Felten D, Cohen N, eds). Academic Press, San Diego,
and others (1989) in that they cannot be pointed as direct
evidence of supernormal conditioning, because there is Ader R, Cohen N (2001) Psychoneuroimmunology: Condi-
no reliable difference between the supernormal and nor-
tioning and immunity. In: Psychoneuroimmunology
mal conditions. However, our results provide indirect
(Ader R, Felten D, Cohen N, eds.). Academic Press, San
evidence of enhancement since the animals trained in
the supernormal condition differed significantly from Ader R, Cohen N, Bovbjerg D (1982) Conditioned suppres-
animals in the non-conditioned control conditions,
sion of humoral immunity in the rat. J Comp Physiol
whereas the animals in the normal condition did not.
Ader R, Kelly K, Moynihan JA, Grota LJ, Cohen N (1993)
Conditioned enhancement of antibody production usingantigen as the unconditioned stimulus. Brain Behav
In conclusion, the results of the present set of experi-
ments provide further evidence that the immune system Alvarez-Borda B, Ramírez-Amaya V, Pérez-Montfort R,
can be associatively conditioned by Pavlovian proce-
Bermúdez-Rattoni F (1995) Enhancement of antibody
dures. The small size of the effect has been a recurrent
production by a learning paradigm. Neurobiol Learn
finding in this area of research, and here it is shown that
it can be enhanced. Further studies may explore other Bovbjerg D, Ader R, Cohen N (1984) Acquisition and
phenomena of Pavlovian conditioning that may influ-
extinction of conditioned suppression of a graft-vs-host
ence the strength of the conditioned response. Examples
response in the rat. J Immunol 132: 111113.
of this are the so-called timing effects, in which the Cohen J (1973) Eta-squared and partial eta-squared in fixed
temporal relationship among stimuli appears to play
factor ANOVA designs. Educational and Psychological
a role in the strength of conditioning. Among the many
temporal phenomena that have been described, one of Cohen N, Moynihan JA, Ader R (1994) Pavlovian condi-
the simplest is the observation that there is an optimal
tioning of the immune system. Int Arch Allerg Immunol
CSUS interval. This phenomenon has been demon-
strated in virtually all Pavlovian conditioning prepara- Dickinson A (1977) Appetitive-aversive interactions:
tions (see: Vogel et al. 2006) but has not been examined
Superconditioning of fear by an appetitive CS. Q J Exp
in conditioned immunomodulation procedures.
Dunn AJ (1989) Psychoneuroimmunology for the psy-
choneuroendocrinologist a review of animal studies ofnervous system-immune system interactions. Psycho-
This research was supported by Fondecyt Grant Nr.
Exton MS, Von Auer AK, Buske-Kirschbaum A, Stockhorst
U, Gobel U, Schedlowski M (2000) Pavlovian condition-
ing of immune function: animal investigation and thechallenge of human application. Behav Brain Res 110:
Ader R (1981) A historical account of conditioned immuno-
biologic responses. In: Psychoneuroimmunology (Ader
Gorczynski R M, Macrae S, Kennedy M (1982) Conditioned
R, ed.). Academic Press, New York, p. 321352.
immune response associated with allogenic skin grafts in
Ader R (2003) Conditioned immunomodulation: Research
needs and directions. Brain Behav Immun 17: 5157.
Hucklebridge F (2002) Behavioral conditioning of the
Ader R, Cohen N (1975) Behaviorally conditioned immuno-
immune system. Int Rev Neurobiol 52: 325351.
suppression. Psychosom Med 37: 333340.
Krank MD, McQueen GM (1988) Conditioned compensa-
Ader R, Cohen N (1982) Behaviorally conditioned immuno-
tory responses elicited by environmental signals for
suppression and murine systemic lupus erythematosus.
cyclophosphamide-induced suppression of the immune
Enhancement of conditioned inmunosuppression 81
Kusnecov A, King MG, Husband AJ (1989) Immunomodu-
Rogers MP, Reich P, Strom TB, Carpenter CB (1976)
lation by behavioral conditioning. Biol Psychol 28:
Behaviorally conditioned immunosuppression: Replica-
tion of recent study. Psychom Med 38: 447451.
Madden KS, Boehm GW, Lee SC, Grota LJ, Cohen N, Ader
Solvason HB, Ghanta VK, Lorden JF, Soong SJ, Hiramoto
R (2001) One-trial conditioning of the antibody response
RN (1991) A behavioral augmentation of natural immu-
to hen egg lysozyme in rats. J Neuroimmunol 113:
nity: Odor specificity supports a Pavlovian conditioning
McQueen GM, Siegel S (1989) Conditional immunomodu-
Stockhorst U, Klosterhalfen S (2005) Conditioning mecha-
lation following training with cyclophosphamide. Behav
nisms and psychoneuroimmunology. Psychother Psycho-
Navarro JI, Hallam SC, Matzel LD, Miller RR (1989)
Vogel EH, Soto FA, Castro ME, Solar PA (2006)
Superconditioning and overshadowing. Learn Motiv 20:
Mathematical models of classical conditioning: Evolution
and current challenges. Rev Latinoam Psicol 38: 215243.
OReilly CA, Exon JH (1986) Cyclophosphamide-Condi-
Wagner AR (1971) Elementary associations. In: Essays in
tioned Suppression Of The Natural-Killer-Cell Response
Neobehaviorism (Kendler HH, Spence JT, eds).
In Rats. Physiol Behav 37: 759764.
AppletonCenturyCrofts, New York, p. 187213.
Rescorla RA (1971) Variation in the effectiveness of rein-
Wayner EA, Flannery GR, Singer G (1978) Effects of taste
forcement and nonreinforcement following prior inhibito-
aversion conditioning on the primary antibody response
ry conditioning. Learn Motiv 2: 113123.
to sheep red blood cells and Bruceella abortus in the albi-
Rescorla RA (2004) Superconditioning from a reduced rein-
no rat. Physiol Behav 21: 9951000.
forcer. Q J Exp Psychol 57B: 133152.
Williams DA, Mehta R, Dumont JL (2004) Conditions
Rescorla RA, Wagner AR (1972) A theory of Pavlovian
favoring superconditioning of irrelevant conditioned
conditioning: Variations in the effectiveness of rein-
stimuli. J Exp Psychol Anim Behav Process 30: 148159.
forcement and nonreinforcement. In: Classical condi-
Williams BA, McDevitt MA (2002) Inhibition and super-
tioning II: Current theory and research (Black AH,
Prokasy WF, eds). AppletonCenturyCrofts, NewYork, p. 6499. Received 22 December 2006, accepted 16 February 2007
PRIMA DELL’USO LEGGETE CON ATTENZIONE TUTTE LE INFORMAZIONI CONTENUTE NEL FOGLIO ILLUSTRATIVO Questo è un medicinale di AUTOMEDICAZIONE che può essere usato per curare disturbi lievi e transitori facilmente riconoscibili e risolvibili senza ricorrere all’aiuto del medico. Può essere quindi acquistato senza ricetta ma va usato cor- rettamente per assicurarne l’efficacia e ridurre gli eff
 Enhancement of conditioned inmunosuppression 77
The main effect of group was explored further
through nine planned contrasts. The first three contrasts
indicate that there was no reliable difference between
groups paired-high and unpaired-high (P=0.784, partial
h2=0.001); between groups paired-medium and
unpaired-medium (P=0.876, partial h2=0.000); or
between groups paired-low and unpaired-low (P=0.626,
partial h2=0.003). On the other hand, the mean antibody
titers of group vehicle was significantly greater than that
of every other group (Ps<0.07, partial h2s>0.077), indi-
cating a residual effect of CY. Thus, contrary to what
was observed with CTA, there was no evidence of con-
ditioned immunomodulation in this experiment.
Enhancement of conditioned inmunosuppression 77
The main effect of group was explored further
through nine planned contrasts. The first three contrasts
indicate that there was no reliable difference between
groups paired-high and unpaired-high (P=0.784, partial
h2=0.001); between groups paired-medium and
unpaired-medium (P=0.876, partial h2=0.000); or
between groups paired-low and unpaired-low (P=0.626,
partial h2=0.003). On the other hand, the mean antibody
titers of group vehicle was significantly greater than that
of every other group (Ps<0.07, partial h2s>0.077), indi-
cating a residual effect of CY. Thus, contrary to what
was observed with CTA, there was no evidence of con-
ditioned immunomodulation in this experiment.
 tive unpaired group. This latter analysis revealed a reli-
able conditioning effect in group paired-three trials,
whose mean immune response was significantly lower
than the unpaired-three trials group (P=0.038, partial
h2=0.103). Groups paired one-trial and unpaired one-
trial did not differ reliably (P=0.648, partial h2=0.005).
tive unpaired group. This latter analysis revealed a reli-
able conditioning effect in group paired-three trials,
whose mean immune response was significantly lower
than the unpaired-three trials group (P=0.038, partial
h2=0.103). Groups paired one-trial and unpaired one-
trial did not differ reliably (P=0.648, partial h2=0.005).