Die Struktur von Tadalafil erlaubt eine selektive Bindung an die Bindungsstelle der PDE5 und minimiert gleichzeitig die Interaktion mit PDE6, was visuelle Nebenwirkungen einschränkt. Seine Verteilung im Organismus erfolgt breit, wobei das Verteilungsvolumen etwa 63 Liter beträgt. Über 90 % des Wirkstoffs sind an Plasmaproteine gebunden. Die Wirkung bleibt unabhängig von der Nahrungsaufnahme konstant. Der Abbauweg über CYP3A4 kann durch Hemmer wie Ritonavir oder Ketoconazol verlangsamt werden, was die Plasmakonzentrationen deutlich erhöht. In diesem Kontext wird cialis 20mg preis häufig in Bezug auf pharmakokinetische Wechselwirkungen erwähnt.
Jid.oxfordjournals.org
Reemergence of Chloroquine-SensitivePlasmodium falciparum Malaria after Cessationof Chloroquine Use in Malawi
James G. Kublin,1,3,a,b Joseph F. Cortese,1,a Eric Mbindo Njunju,5 Rabia A. G. Mukadam,3 Jack J. Wirima,3 Peter N. Kazembe,4 Abdoulaye A. Djimde´,6 Bourema Kouriba,6 Terrie E. Taylor,2,3 and Christopher V. Plowe1
1Malaria Section, Center for Vaccine Development, University of Maryland School of Medicine, Baltimore; 2Department of Internal Medicine,College of Osteopathic Medicine, East Lansing, Michigan; 3Blantyre Malaria Project, College of Medicine, University of Malawi, Blantyre,and 4Lilongwe Central Hospital, Lilongwe, Malawi; 5Tropical Diseases Research Centre, Ndola, Zambia; 6Malaria Research and Training Center,
In 1993, Malawi became the first African country to replace chloroquine with sulfadoxine-pyrimethamine nationwide in response to high rates of chloroquine-resistant falciparum malaria. To determine whether withdrawal of chloroquine can lead to the reemergence of chloroquine sensitivity, the prevalence of the pfcrt 76T molecular marker for chloroquine-resistant Plasmodium falciparum malaria was retrospectively measured in Blantyre, Malawi. The prevalence of the chloroquine-resistant pfcrt genotype decreased from 85% in 1992 to 13% in 2000. In 2001, chloroquine cleared 100% of 63 asymptomatic P. falciparum infections, no isolates were resistant to chloroquine in vitro, and no infections with the chloroquine-resistant pfcrt genotype were detected. A concerted national effort to withdraw chloroquine from use has been followed by a return of chloroquine-sensitive falciparum malaria in Malawi. The reintroduction of chloroquine, ideally in combination with another antimalarial drug, should be considered in areas where chloroquine resistance has declined and safe and affordable alternatives remain unavailable.
In 1993, Malawi became the first sub-Saharan African
(SP) to the antimalarial of first choice nationwide. The
country to discontinue the routine use of chloroquine
decision was based on chloroquine’s increasing failure to
against Plasmodium falciparum malaria and to elevate
produce adequate clinical and hematological recovery.
the antifolate combination sulfadoxine-pyrimethamine
High-level parasitological resistance was observed in
180% of Malawian children treated with chloroquine in
1990 [1]. Since 1993, SP has been the only available
Received 25 November 2002; accepted 27 January 2003; electronically published
treatment for uncomplicated malaria in all government
health facilities and is dispensed without prescription.
Presented in part: 51st annual meeting of the American Society of Tropical
Medicine and Hygiene, Denver, 10–14 November 2002 (abstract 54).
Although chloroquine has remained available by pre-
Informed consent was obtained from subjects or their parents or guardians,
scription and through unauthorized private sources, a
according to the human experimentation guidelines of the US Department of Health
national information campaign was largely successful in
and Human Services and/or those of the authors’ institutions.
Financial support: National Institute of Allergy and Infectious Diseases (grant
convincing health practitioners and the public to accept
SP as the treatment of choice for children with malaria.
a J.G.K. and J.F.C. contributed equally to this work.
Other African countries, including Kenya in 1999 [2],
b Present affiliation: Merck Research Laboratories, West Point, Pennsylvania.
have instituted similar changes in national drug policy
Reprints or correspondence: Dr. Christopher V. Plowe, Malaria Section, Center
for Vaccine Development, University of Maryland at Baltimore, 685 W. Baltimore
in the face of increasing chloroquine resistance.
St., HSF 480, Baltimore, MD ([email protected]).
Chloroquine resistance in P. falciparum is conferred
The Journal of Infectious Diseases 2003; 187:1870–5
by mutations in the parasite pfcrt, which encodes a
ᮊ 2003 by the Infectious Diseases Society of America. All rights reserved. 0022-1899/2003/18712-0006$15.00
putative transporter localized to the digestive vacuole
1870 • JID 2003:187 (15 June) • Kublin et al.
[3, 4]. One mutation, K76T, was perfectly associated with in
.umaryland.edu/CVD/plowe.html. In brief, the dried thick smear
vitro resistance in all progeny of a genetic cross between chlo-
was detached from the slide with a new razor blade, and the
roquine-sensitive and -resistant parental clones and among a
debris were collected in a microcentrifuge tube. DNA was pre-
set of geographically diverse parasite isolates [3]. Several field
pared for PCR by a methanol-fixation/heat extraction method,
studies summarized in a recent review have since confirmed
as described elsewhere [7], for preparation of DNA from dried
the absolute specificity of the pfcrt K76T to clinical chloroquine
blood spots. Nested PCR and subsequent allele-specific restric-
resistance [5]. Polymorphisms in pfmdr1, encoding the P. fal-
tion analysis were performed to identify polymorphic codons of
ciparum P glycoprotein homologue 1, modulate chloroquine
interest. Infections presenting with mixed alleles in any assay
resistance in mutant pfcrt-harboring parasites in vitro [6], al-
were defined as polyclonal and were characterized as mutant for
though their role in vivo has yet to be substantiated [7]. SP
that assay. Direct DNA sequencing was performed at the Uni-
resistance is conferred by mutations in 2 genes encoding en-
versity of Maryland Baltimore Biopolymer Facility. The x2 test
zymes in the parasite’s folate synthesis pathway, dihydrofolate
for linear trend using Epi Info (version 6.04c; CDC) was used
reductase (dhfr) and dihydropteroate synthase [8].
It is not known whether years of reliance on antimalarials
In vitro chloroquine assays.
other than chloroquine can lead to the reemergence of chloro-
collected in February 2001 at the Ndirande Health Centre from
quine-sensitive P. falciparum and permit the reintroduction of
consenting individuals presenting with uncomplicated malaria.
this safe and affordable drug. Using polymerase chain reaction
Drug-sensitivity assays were performed essentially, as described
(PCR)–based assays, we measured the prevalence of resistance-
elsewhere [10], using the World Health Organization (WHO)
conferring mutations in malaria-infected human blood samples
microtest method, a field test that estimates drug susceptibility
obtained from southern Malawi before and after the 1993 change
by culturing fresh parasites for 1 48-h life cycle in the presence
in drug policy. We demonstrate a progressive decline in the fre-
of increasing drug concentrations and by determining parasite
quency of the pfcrt K76T mutation over the decade after the
survival by light microscopy. In brief, 0.1 mL of blood was
proscription of chloroquine and a much lower prevalence of 76T
collected in a sterile, 100-mL heparin-treated capillary tube and
in Malawi than in neighboring Zambia, where chloroquine re-
immediately was added to 0.9 mL of RPMI 1640 culture me-
mained in use. We confirm these molecular findings by dem-
dium (Life Technologies). Blood-medium mixture (50 mL) was
onstrating chloroquine sensitivity in vitro and in vivo in con-
added to each well of tissue culture plates predosed with chloro-
temporary infections from this same region of Malawi.
quine diphosphate salt (Sigma) at 1, 2, 4, 8, 16, 32, and 64pmol/well. The plates were incubated at 37.5ЊC for 30 h, ac-cording to standard methodology. After incubation, Field’s-
MATERIALS AND METHODS
stained thick blood films were prepared, and the number of
Study sites and sample sources.
mature schizonts was counted per 200 asexual parasites. Ac-
residents in southern Malawi with moderate-to-intense levels
cording to the WHO protocol, chloroquine susceptibility was
of seasonal P. falciparum transmission that peak during De-
defined as complete schizont inhibition at р4 pmol of chloro-
cember through March. Archived, anonymized microscope
quine, and chloroquine resistance was defined as schizont for-
slides with Field’s-stained thick smears were provided by the
mation at у8 pmol of chloroquine. Isolates that formed schiz-
Malaria Project, Blantyre, Malawi. These slides had been col-
onts at 4 pmol of chloroquine but had complete schizont in-
lected in the January–June malaria seasons in 1992–1996 from
hibition at 8 pmol were considered to be intermediate.
children hospitalized for severe malaria. The 1998–2000 Ma-
In vivo chloroquine efficacy.
lawian samples were collected at the Ndirande Health Centre
vivo chloroquine efficacy studies showed unacceptably high
in Blantyre from children with uncomplicated malaria, ac-
rates of treatment failure, it was deemed unethical to assess
cording to standard definitions [9], and were stored as dried
chloroquine efficacy in persons with symptomatic malaria or
blood spots on filter paper, as described elsewhere [7]. The
in children, who are at higher risk of becoming symptomatic.
Zambian samples were collected in 1999 as filter papers and
Therefore, we conducted a preliminary in vivo chloroquine
blood smears from symptomatic children in the outpatient de-
efficacy study in infected adults with no symptoms. Consenting
partment of the Isoka district hospital in the northern province.
asymptomatic adults accompanying sick children to the Ndi-
All available samples from 1992–1996 and randomly selected
rande Health Centre in January–September 2001 were screened
samples from the Malawi 1998–2000 and Zambia 1999 sample
for occult P. falciparum infection by standard microscopy.
sets were analyzed. All samples were obtained before initiation
Those with positive blood smears were treated by directly ob-
served therapy with standard doses of two 300-mg tablets of
Sample analysis.
chloroquine phosphate on the treatment day and on day 1 after
tion-specific assays are detailed on the Internet: http://medschool
treatment and one 300-mg tablet on day 2 after treatment.
Chloroquine-Sensitive Malaria in Malawi • JID 2003:187 (15 June) • 1871
Chloroquine efficacy was measured after slight modificationsof standard protocols, as described elsewhere [11], with activemicroscopic and clinical follow-up on days 3, 7, and 14 aftertreatment and passive surveillance by continuous availabilityof medical care throughout the follow-up period. Because studyparticipants were asymptomatic, parasitologic rather than ther-apeutic outcome definitions were used; efficacy results wererecorded as sensitive or as resistant at the RI, RII, or RIII levels[11]. An additional follow-up assessment was conducted onday 28 for study participants who could be located at this time.
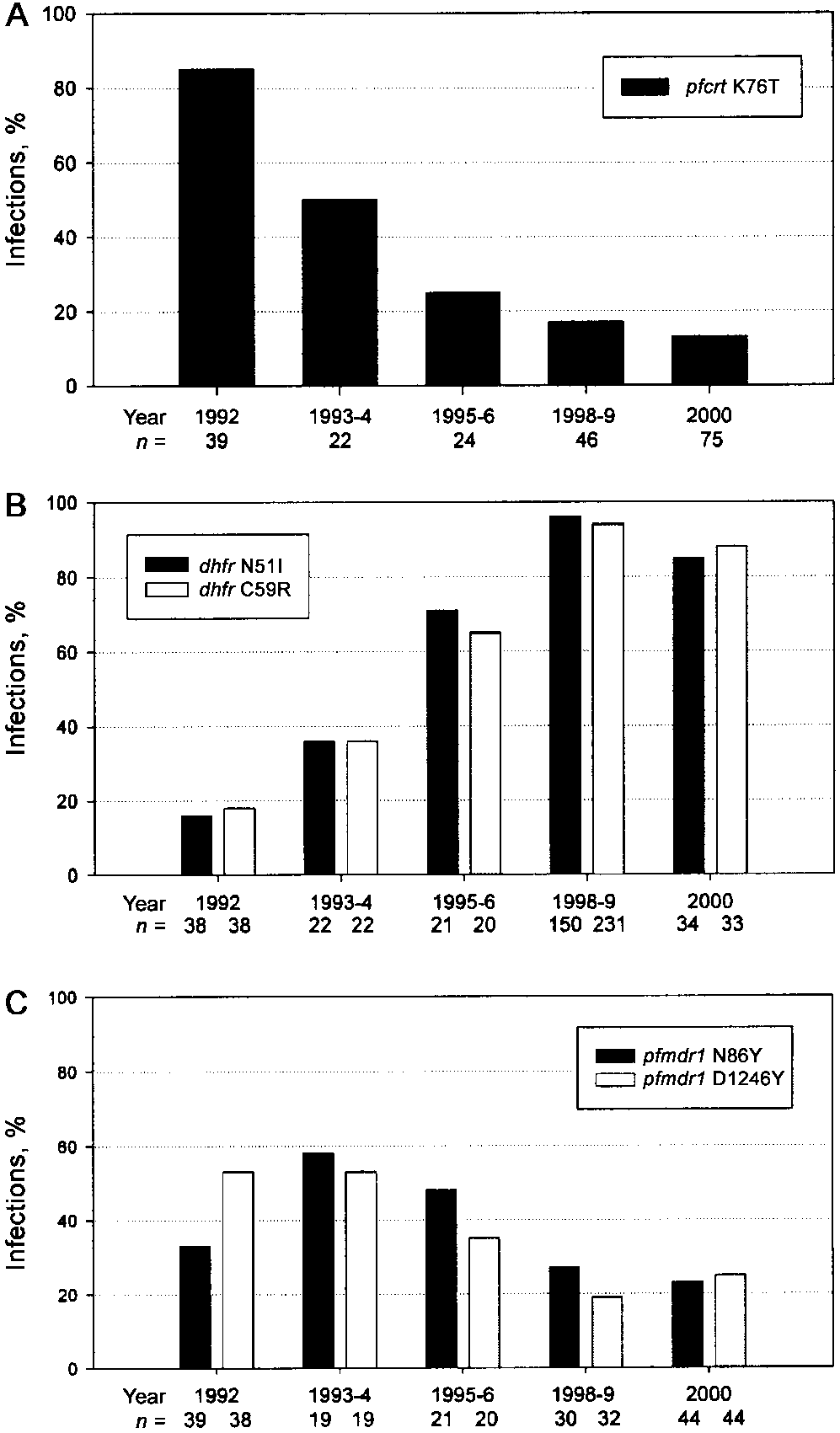
DNA was recovered, and assays for pfcrt 76T were performedsuccessfully on 39/40 thick smears from 1992, 23/24 from 1993–1994, and 24/27 from 1995–1996. Similar results were obtainedfor the other assays (sample sizes are shown in figure 1). Success
rates for 1998–2000 filter-paper samples were even higher, rang-ing from 93% to 100% (data not shown). The number of1998–1999 samples analyzed for dhfr mutations was higher,because a large set of samples had been analyzed previously for
The chloroquine resistance mutation pfcrt 76T was detected
in 30 (85%) of the 39 isolates obtained in 1992 (figure 1A),and its prevalence diminished progressively and significantly(P ! .000 )
1 over the subsequent 8 years, from 11 (50%) of 22
isolates obtained in 1993–1994 to 10 (13%) of 75 isolates in2000. No infections with pfcrt 76T were found among 25 sam-
ples collected in 2001 that were analyzed for both in vivo chlo-roquine resistance and the presence of this mutation. Thecurrent low prevalence of the pfcrt 76T in Malawi is in sharpcontrast to its frequency in neighboring Zambia, where chloro-quine has remained the first-line antimalarial drug. We analyzed
Figure. 1.
Prevalence of drug-resistance mutations in Plasmodium
isolates obtained in 1999 from the Isoka region of northern
falciparum malaria infections in Malawi before and after chloroquine was
Zambia, 80 km from the western border of Malawi, and found
replaced with sulfadoxine-pyrimethamine (SP) in 1993. A, Chloroquine
a 92% prevalence of the mutation (46/50 infections). These
resistance–conferring 76T mutation in pfcrt; B, pyrimethamine resistance–conferring N51I and C59R mutations in dhfr; and C, possible chloroquine
findings in Zambia are similar to those of a recent study con-
resistance–modulating N86Y and D1256Y mutations in pfmdr1. Sample
ducted in southern Mozambique, where a 91% prevalence of
sizes are indicated beneath the years.
the mutant pfcrt was detected [12].
The institution of SP was followed by an increased prevalence
of 2 mutations in the parasite dhfr that confer pyrimethamine
some studies with chloroquine resistance [6, 15] and for which
resistance and are common throughout East and Central Africa
a role in antifolate resistance has not been described. Both
[13]. In 1992, the dhfr N51I and C59R were present in only
mutations exhibited a modest decrease in frequency over the
16 (6%) of 38 and 7 (18%) of 38 Malawi infections, respectively
decade, with the decline in D1246Y, from one-half of the in-
(figure 1B). By 1999, nearly all infections were mutant at these
fections in 1993 to one-fourth in 2000, attaining statistical sig-
codons: N51I, 96% (144/150); and C59R, 94% (217/231). The
significant increase in dhfr mutations (P ! .0001) is consistent
The 76T has invariably been found to be accompanied by
with the increasing rates of antifolate resistance evident in Ma-
sets of additional pfcrt mutations [3]. We analyzed a subset of
the infections from 1993 (8 with 76K infections and 4 with
We found less dramatic trends for 2 mutations in the pfmdr1
76T) and 1998–2000 (10 with 76K infections and 2 with 76T)
(figure 1C), N86Y and D1246Y, which have been associated in
for the resistance-associated mutations at pfcrt codons 74, 75,
1872 • JID 2003:187 (15 June) • Kublin et al.
220, 271, and 371 and found that the full array of mutations
in selection of drug-sensitive phenotypes [17, 18]. Incomplete
always cosegregated with 76T, whereas none of the infections
withdrawal of the drug, cross-resistance to agents still in use,
that were exclusively 76K were mutant at these codons (data
and compensatory mutations that reduce the burden of resis-
not shown). Thus the pfcrt in Malawi appears to have main-
tance without jeopardizing the resistance itself [19] may allow
tained the integrity of its fully mutant and wild alleles, and the
for resistant organisms to persist, despite restricted availability
reemerging 76T pfcrt are not mutants that simply reverted to
of an established antimicrobial agent. In P. falciparum malaria
K76, which alone can confer the in vitro chloroquine-sensitive
in Malawi, the in vivo fitness burden of harboring the mutant
phenotype [3]. Variation was detected at pfcrt codons 326 and
pfcrt appears to be substantial in the absence of selective drug
356, which are polymorphic but not associated with resistance
pressure, and with the mechanisms of antifolate resistance dis-
[7], among both the sensitive and the resistant infections (data
tinct from those of the aminoquinolines, there appears to have
not shown). This heterogeneity at the pfcrt locus, along with
been a strong selection of chloroquine-sensitive parasites after
our finding that 55% of pfcrt 76T-harboring infections over the
the nationwide replacement of chloroquine with SP.
time course were mutant at dhfr codons 51 and/or 59 (data
The slower decline in prevalence of mutations in pfmdr1 sug-
not shown), are indicative of the polyclonal nature of malaria
gests that these mutations may be less deleterious to parasite
infections in this setting and the high recombination rates of
fitness than are pfcrt mutations. Although genetic transformation
P. falciparum in Africa [16]. Malaria harboring the sensitive
experiments have found that pfmdr1 mutations can modulate
pfcrt appears to have arisen from low levels in the indigenous
the level of chloroquine resistance once it has been conferred by
parasite populations and/or to have emigrated from bordering
mutations in pfcrt, mutations in pfmdr1 are by themselves in-
countries where chloroquine pressure persists. But we can ex-
sufficient to confer chloroquine resistance [20]. Recent studies
clude as an explanation for the diminished prevalence of the
have reported that pfmdr1 mutations, in addition to pfcrt mu-
mutant pfcrt a selective clonal replacement of antifolate-sensi-
tations, are no more strongly associated with chloroquine treat-
tive/chloroquine-resistant parasites by antifolate-resistant para-
ment failure than pfcrt mutations alone [7] and that pfmdr1
sites that coincidentally harbor a sensitive pfcrt.
mutations do not add to the predictive value of pfcrt mutations
Of the 11 isolates successfully assayed for in vitro drug suscep-
for chloroquine treatment failure [21]. Even if pfmdr1 mutations
tibility, 9 (82%) were determined to be fully chloroquine-sensitive,
do contribute to treatment failure, they require pfcrt mutations
whereas 2 isolates were of intermediate chloroquine susceptibility.
to exert an effect on the response to chloroquine. Therefore,
No isolates exhibited in vitro chloroquine resistance.
persistence of pfmdr1 mutations in a population with a very low
Seventy-two adults were enrolled in the study of in vivo
prevalence of pfcrt mutations would not be expected to reduce
chloroquine resistance. As expected, initial parasitemias were
chloroquine efficacy in that population.
low, with a geometric mean of 600 asexual forms/mm3 and a
The increase in mutations in dhfr is consistent with what is
range of 50–12,250 asexual forms/mm3. Nine adults were lost
known about SP efficacy in this area during the 1990s. Before
to follow-up before day 3, precluding any determination of
SP was introduced in Malawi as the first-line drug, efficacy
outcome. Each of 63 remaining asymptomatic P. falciparum
rates as high as 100% were reported [22], and parasitological
infections in adults was successfully cleared within 3 days after
cure rates of 90.5% were found in southern Malawi 1 year after
chloroquine treatment without development of fever or other
the introduction of SP [23]. Our own studies from the site of
symptoms of malaria. Fifty-five adults completed the 14-day
the current study have found that, by 2000, SP’s parasitological
follow-up, and 42 were followed-up for 28 days. Five partici-
cure rate had decreased to !70% and its clinical efficacy to
pants were lost to follow-up after treatment day 3, and 3 were
∼80% (authors’ unpublished data), corresponding to the in-
lost to follow-up after day 7. No recurrent infections were
creasing prevalence of dhfr mutations.
detected during the follow-up period; thus, 100% had sensitive
Standard oral doses of chloroquine cleared asymptomatic P.falciparum infections in adults with no exceptions, and therewas no evidence of recurrent infection among the 53 subjects
DISCUSSION
monitored for at least 14 days, thus meeting the criteria forsensitive parsitological outcome in high transmission areas.
Our findings suggest that chloroquine-sensitive P. falciparum
Some loss to follow-up was expected in this asymptomatic
parasites have reemerged and are now predominant at a site
urban adult population, and we cannot rule out the possibility
in Africa where chloroquine was withdrawn in 1993, whereas
of recurrent infections among subjects who were lost to follow-
high levels of chloroquine resistance have persisted unabated
up between 3 and 14 days after treatment. However, the com-
in nearby areas where the drug has remained in use. The with-
plete lack of recurrent infections in the 76% of subjects who
drawal of antimicrobial drug pressure does not always com-
were followed for у14 days is strong evidence of chloroquine’s
promise the fitness of resistant microorganisms and result
ability to clear P. falciparum infections in this setting.
Chloroquine-Sensitive Malaria in Malawi • JID 2003:187 (15 June) • 1873
It is possible that some of these semi-immune adults would
other drugs that will eventually fail. Many African countries
have spontaneously cleared their malaria infections even with-
are delaying decisions to change antimalarial therapies, despite
out drug treatment, but it was felt to be unethical to include
high rates of chloroquine resistance that are associated with
a placebo or nontreatment arm in this first in vivo assessment
increasing malaria-attributable disease and death, fearing a slip-
of chloroquine in Malawi since the drug was withdrawn. On
pery slope of increasingly expensive antimalarial drugs with
the basis of the preliminary evidence of chloroquine efficacy
limited useful therapeutic lives. The preliminary in vivo data
presented here, a controlled trial of chloroquine alone or in
reported here justify the conduct of controlled trials of chlo-
combination with other drugs may now be ethically justifiable
roquine efficacy in areas where chloroquine use has been sub-
stantially reduced for a period of years. If these trials confirm
Nine of 11 fresh P. falciparum isolates were sensitive to chlo-
a return of chloroquine’s clinical efficacy, governments can con-
roquine in vitro, consistent with a recent report of a high prev-
sider withdrawing chloroquine and switching to other drugs
alence of in vitro chloroquine-sensitive P. falciparum in central
on an interim basis, knowing that they may be able to later
Malawi [24]. The finding that 2 infections yielded microtest val-
reintroduce chloroquine, which is unparalleled in its safety and
ues in the intermediate range is most likely explained by inac-
low cost. However, it must be emphasized that Malawi made
curacies of the in vitro test, or by mixed resistant and sensitive
a concerted and sustained effort to withdraw chloroquine from
parasites infections. Microtest assays are well known to be sub-
use and that less rigorous efforts to reduce chloroquine drug
ject to variation and artifact, and, although they are useful as
pressure may not result in the same reemergence of chloroquine
general measures of in vitro resistance in field surveys, they lack
the reliability to definitively characterize individual infections
Chloroquine’s useful therapeutic life span in Malawi is al-
[25–27]. P. falciparum infections in settings like Malawi also
most certain to be short-lived if it is reintroduced as mono-
are frequently polyclonal, so that discrepant results could arise
therapy. There is growing consensus that combination chemo-
if in vitro survival of a minor number of resistant parasites is
therapy should be the rule in malaria treatment, to deter the
detected among a predominantly sensitive population. Such par-
development of drug resistance, and that the artemesinin de-
asites would have had to be too few to be detected by the mo-
rivatives, because of their rapid reduction of parasite biomass
lecular assays, which may not detect subpopulations comprising
and continued efficacy against multidrug resistant P. falciparum,should constitute a component of most such regimens [34].
!5%–10% of an infection. A more careful evaluation of the
chloroquine in vitro response using labeled nutrient uptake as-
As SP failure rates increase, the reintroduction of chloroquinein Malawi in combination with artesunate or with other short-
says and assessing the verapamil-reversal phenotype associated
acting drugs, such as chlorproguanil-dapsone, should be con-
with chloroquine resistance would have been more definitive but
sidered and would likely extend the useful life span of each of
was not possible on these fresh isolates collected under field
the drugs. This study reinforces the need for rational anti-
malarial drug policies at both the national and regional levels
Earlier reports from China [28] and Gabon [29] showed less
and demonstrates the utility of surveillance using molecular
dramatic declines in measures of chloroquine resistance after
assays to monitor the effects of these policies on drug resistance.
reductions in chloroquine use. However, regions of Columbiaand Venezuela, where chloroquine use is reportedly minimized,continue to sustain mutant pfcrt 76T–harboring parasites [30],
Acknowledgments
which suggests that the trends observed in Malawi may not bereplicated in settings with different epidemiological and trans-
We thank Standwell Nkhoma, Jean-Claude Akpa, and Love-
mission characteristics. Nonetheless, in malaria-endemic coun-
more Kaetano, for technical assistance, and Rosemary Musonda,
tries with semi-immune host populations, such as Malawi, even
a partial resumption of chloroquine sensitivity may positivelyimpact public health. In Mali, up to two-thirds of infectionscarrying the mutant pfcrt are cured by chloroquine treatment,
References
an outcome due, at least in part, to the contribution of host
1. Bloland PB, Lackritz EM, Kazembe PN, Were JB, Steketee R, Campbell
immunity to parasite clearance [31, 32]. By inference, the 13%
CC. Beyond chloroquine: implications of drug resistance for evaluating malaria therapy efficacy and treatment policy in Africa. J Infect Dis 1993;
prevalence of mutant pfcrt seen in 2000 in Malawi predicts a
chloroquine failure rate !5%, although chloroquine failure rates
2. Shretta R, Omumbo J, Rapuoda B, Snow RW. Using evidence to change
150% have recently been reported in both Zambia [33] and
antimalarial drug policy in Kenya. Trop Med Int Health 2000; 5:755–64.
3. Fidock DA, Nomura T, Talley AK, et al. Mutations in the P. falciparum
digestive vacuole transmembrane protein PfCRT and evidence for their
Our results support the possibility of reintroducing chloro-
role in chloroquine resistance. Mol Cell 2000; 6:861–71.
quine to areas where it has been withdrawn and replaced by
4. Sidhu AB, Verdier-Pinard D, Fidock DA. Chloroquine resistance in
1874 • JID 2003:187 (15 June) • Kublin et al. Plasmodium falciparum malaria parasites conferred by pfcrt mutations.
pression in isoniazid-resistant Mycobacterium tuberculosis. Science1996; 272:
Science 2002; 298:210–3.
5. Wellems TE, Plowe CV. Chloroquine-resistant malaria. J Infect Dis 2001;
20. Reed MB, Saliba KJ, Caruana SR, Kirk K, Cowman AF. Pgh1 modulates
sensitivity and resistance to multiple antimalarials in Plasmodium falci-
6. Reed MB, Saliba KJ, Caruana SR, Kirk K, Cowman AF. Pgh1 modulates
parum. Nature 2000; 403:906–9.
sensitivity and resistance to multiple antimalarials in Plasmodium fal-
21. Jelinek T, Aida AO, Peyerl-Hoffmann G, et al. Diagnostic value of mo-
ciparum. Nature 2000; 403:906–9.
lecular markers in chloroquine-resistant falciparum malaria in Southern
7. Djimde´ A, Doumbo OK, Cortese JF, et al. A molecular marker for chlo-
Mauritania. Am J Trop Med Hyg 2002; 67:449–53.
roquine-resistant falciparum malaria. N Engl J Med 2001; 344:257–63.
22. Heymann DL, Khoromana CO, Wirima JJ, Campbell CC. Comparative
8. Plowe CV, Kublin JG, Doumbo OK. P. falciparum dihydrofolate reductase
efficacy of alternative primary therapies for Plasmodium falciparum
and dihydropteroate synthase mutations: epidemiology and role in clinical
infections in Malawi. Trans R Soc Trop Med Hyg 1987; 81:722–4.
resistance to antifolates. Drug Resist Updats 1998; 1:389–96.
23. Verhoeff FH, Brabin BJ, Masache P, Kachale B, Kazembe P, van der
9. World Health Organization. Assessment of therapeutic efficacy of antima-
Kaay HJ. Parasitological and haematological responses to treatment of
larial drugs for uncomplicated falciparum malaria in areas with intense
Plasmodium falciparum malaria with sulphadoxine-pyrimethamine in
transmission: Report WHO/MAL/96–1077. Geneva: World Health Or-
southern Malawi. Ann Trop Med Parasitol 1997; 91:133–40.
ganization, 1996.
24. Takechi M, Matsuo M, Ziba C, et al. Therapeutic efficacy of sulpha-
10. World Health Organization. In vitro micro-test (Mark III) for the as-
doxine/pyrimethamine and susceptibility in vitro of P. falciparum iso-
sessment of the response of Plasmodium falciparum to chloroquine, me-
lates to sulphadoxine-pyremethamine and other antimalarial drugs in
floquine, quinine, sulfadoxine/pyrimethamine and amodiaquine. Report
Malawian children. Trop Med Int Health 2001; 6:429–34.
no. MAP/87.2. Geneva: World Health Organization, 1987.
25. Wellems TE, Plowe CV. Chloroquine-resistant malaria. J Infect Dis 2001;
11. Plowe CV, Doumbo OK, Djimde A, et al. Chloroquine treatment of
uncomplicated Plasmodium falciparum malaria in Mali: parasitologic re-
26. Durand R, Jafari S, Vauzelle J, Delabre J, Jesic Z, Le Bras J. Analysis
sistance versus therapeutic efficacy. Am J Trop Med Hyg 2001; 64:242–6.
of pfcrt point mutations and chloroquine susceptibility in isolates of
12. Mayor AG, Gomez-Olive X, Aponte JJ, et al. Prevalence of the K76T
Plasmodium falciparum. Mol Biochem Parasitol 2001; 114:95–102.
mutation in the putative Plasmodium falciparum chloroquine resistance
27. Carlin JM, Vande Waa JA, Jensen JB, Akood MA. African serum in-
transporter (pfcrt) gene and its relation to chloroquine resistance in
terference in the determination of chloroquine sensitivity in Plasmo-
Mozambique. J Infect Dis 2001; 183:1413–6. dium falciparum. Z Parasitenkd 1984; 70:589–97.
13. Plowe CV, Cortese JF, Djimde´ A, et al. Mutations in Plasmodium fal-
28. Liu DQ, Liu RJ, Ren DX, et al. Changes in the resistance of Plasmodiumciparum dihydrofolate reductase and dihydropteroate synthase and ep-
falciparum to chloroquine in Hainan, China. Bull World Health Organ
idemiologic patterns of pyrimethamine- sulfadoxine use and resistance. 1995; 73:483–6.
J Infect Dis 1997; 176:1590–6.
29. Schwenke A, Brandts C, Philipps J, Winkler S, Wernsdorfer WH, Kremsner
14. Kublin JG, Dzinjalamala FK, Kamwendo DD, et al. Molecular markers
PG. Declining chloroquine resistance of Plasmodium falciparum in Lam-
for failure of sulfadoxine-pyrimethamine and chlorproguanil-dapsone
barene, Gabon from 1992 to 1998. Wien Klin Wochenschr 2001; 113:63–4.
treatment of Plasmodium falciparum malaria. J Infect Dis 2002; 185:380–8.
15. Duraisingh MT, Drakeley CJ, Muller O, et al. Evidence for selection
30. Cortese JF, Caraballo A, Contreras CE, Plowe CV. Origin and dissem-
for the tyrosine-86 allele of the pfmdr1 gene of Plasmodium falciparum
ination of Plasmodium falciparum drug-resistance mutations in South
by chloroquine and amodiaquine. Parasitology 1997; 114:205–11.
America. J Infect Dis 2002; 186:999–1006.
16. Conway DJ, Roper C, Oduola AM, et al. High recombination rate in
31. Djimde A, Doumbo OK, Steketee RW, Plowe CV. Application of a
natural populations of Plasmodium falciparum. Proc Natl Acad Sci USA
molecular marker for surveillance of chloroquine-resistant falciparum
1999; 96:4506–11.
malaria. Lancet 2001; 358:890–1.
17. Levin BR, Perrot V, Walker N. Compensatory mutations, antibiotic
32. Wellems TE, Plowe CV. Chloroquine-resistant malaria. J Infect Dis 2001;
resistance and the population genetics of adaptive evolution in bacteria.
Genetics 2000; 154:985–97.
33. Bijl HM, Kager J, Koetsier DW, van der Werf TS. Chloroquine- and
18. Enne VI, Livermore DM, Stephens P, Hall LM. Persistence of sulphon-
sulfadoxine-pyrimethamine–resistant falciparum malaria in vivo: a pi-
amide resistance in Escherichia coli in the UK despite national prescribing
lot study in rural Zambia. Trop Med Int Health 2000; 5:692–5.
restriction. Lancet 2001; 357:1325–8.
34. White NJ, Nosten F, Looareesuwan S, et al. Averting a malaria disaster.
19. Sherman DR, Mdluli K, Hickey MJ, et al. Compensatory ahpC gene ex-
Lancet 1999; 353:1965–7.
Chloroquine-Sensitive Malaria in Malawi • JID 2003:187 (15 June) • 1875
PSEUDOMONAS AERUGINOSA SU LARINDA ANT B YOT K D RENC VE METALLO-BETA-LAKTAMAZ SIKLI I * I ıl F DAN, Feryal ÇET N GÜREL K, Sevgi YÜKSEL, Nedim SULTAN * Gazi Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalı, ANKARA Pseudomonas aeruginosa hastane infeksiyonuna neden olan önemli bir etkendir ve çoklu direnç gösteren izolat sıklı ı giderek artmaktadır. Bu �
PDSG newsletter Welcome to this month’s Pick’s Disease Support Group Newsletter • Peter Moore tells us all about the • Events Diary - make sure you book February 2008 • Duncan Hutchison updates us on • Julian Taylor & Margaret Rees www.pdsg.org.uk ANNUAL SEMINAR PICK’S DISEASE SUPPORT FRONTOTEMPORAL DEMENTIA (including Pick’s disease) ST
 Chloroquine efficacy was measured after slight modificationsof standard protocols, as described elsewhere [11], with activemicroscopic and clinical follow-up on days 3, 7, and 14 aftertreatment and passive surveillance by continuous availabilityof medical care throughout the follow-up period. Because studyparticipants were asymptomatic, parasitologic rather than ther-apeutic outcome definitions were used; efficacy results wererecorded as sensitive or as resistant at the RI, RII, or RIII levels[11]. An additional follow-up assessment was conducted onday 28 for study participants who could be located at this time.
Chloroquine efficacy was measured after slight modificationsof standard protocols, as described elsewhere [11], with activemicroscopic and clinical follow-up on days 3, 7, and 14 aftertreatment and passive surveillance by continuous availabilityof medical care throughout the follow-up period. Because studyparticipants were asymptomatic, parasitologic rather than ther-apeutic outcome definitions were used; efficacy results wererecorded as sensitive or as resistant at the RI, RII, or RIII levels[11]. An additional follow-up assessment was conducted onday 28 for study participants who could be located at this time.